

2023年1月17日,由华南农业大学联合中国农业科学院生物技术研究所等多个单位研究成果“De novo genome assembly and analyses of 12 founder inbred lines provide insights into maize heterosis”在线刊登国际知名期刊Nature Genetics!该研究完成了12个玉米基因组的构建,并通过泛基因组、eQTL等手段解析了玉米的杂种优势机制,为加速玉米改良育种提供了重要理论基础和遗传资源。
中国农业科学院生物技术研究所研究员王宝宝、博士研究生候美、中国农业大学已毕业博士史俊鹏(现为中山大学副教授)、河南农业大学教授库丽霞、北京市农林科学院研究员宋伟及中国农业科学院作科所副研究员李春辉为论文共同第一作者,通讯作者为华南农业大学王海洋教授、中国农业大学赖锦盛教授、北京市农林科学院赵久然研究员和河南农业大学陈彦惠教授。百迈客以共同作者参与本项研究。
研究背景
杂种优势是指两个遗传组成不同的亲本杂交产生的杂种F1,在产量、生物量和抗逆性等方面表现优于亲本的现象。玉米约占全球谷物总产量的36%,中国和美国是世界两大玉米主产国,生产量占全球约60%。玉米也因其异交习性、丰富的遗传变异和农业重要性而成为杂种优势研究的模式植物。前期研究致力于剖析玉米杂种优势的机制,包括绘制不同杂交组合中不同农艺性状的杂种优势位点,分析基因表达、基因组分化、表观遗传调控和亲本及其杂交后代中的代谢产物等。然而杂种优势的分子机制仍不清楚,限制杂种优势研究的一个主要因素是缺乏有关亲本的高质量参考基因组及其遗传变异的准确信息。
材料方法
Denovo
①被广泛用于温带玉米育种的11个骨干自交系(FIL):A632(代表SS2组)、Ye478和Zheng58(代表中国PA组)、OH43(代表Lancaster 2组),PH207(代表IDT组)、Xu178(代表PB组)、Jing724(代表X组)、Huangzaosi、Chang7-2和Jing92(代表TSPT组)。Dan340源自中国陆地种族LvDa Red Cob(LRC)。
②代表亚热带群的FIL材料S37。
RNA-seq
131个自交系吐丝期的幼穗(6G)
QTL及GWAS分析
①350个温带优良自交系;
②670个Xu178×HuangC杂交的RIL群体;
③14个FILs双列杂交的91个F1S组合。
主要研究结果
1、12个骨干自交系的亲缘关系及基因组组装注释
为了研究玉米杂种优势群形成的遗传基础并探索玉米杂种优势的分子机制,作者重新分析了在美国和中国广泛用于现代玉米育种的350个优良自交系(IL)的群体结构。结果表明,这些材料可分为10个亚组,分别对应于SS(SS1、SS2)、NSS(Lancaster1、Lancaster 2)、IDT、PA、TSPT、PB组、X组和混合组(图1)。并基于系统发育树,选择了12个骨干自交系(FIL)用于高质量的基因组组装和注释。这些自交系中,每一个FIL都曾被用来培育几十或数百个不同杂种群体的优良亲本系,并在现代杂交玉米育种中发挥了关键作用,具有广泛的代表性。
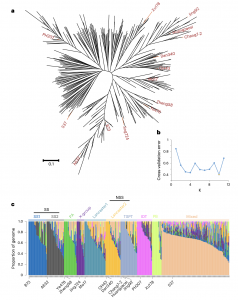
图1 现代玉米育种中典型玉米自交系的群体结构分析
通过三代组装,12个材料组装大小在2.18 Gb~2.31 Gb;Contig N50(0.27 Mb-2.07 Mb)、Scaffold N50(4.36 Mb-15.34 Mb)、染色体挂载率94.3%-97.6%(表1),平均预测了44,554个蛋白质编码基因(42,111~46,320个)。通过二代reads评估(平均~99.26%)、BUSCO评估(基因组平均~96%;基因平均~97.2%)、LAI评估(平均~27.16)等结果显示,本次构建了高质量和高完整性的玉米基因组。
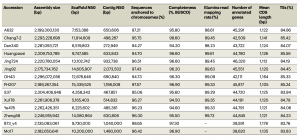
表1 14个玉米材料基因组结果统计
重复元件是玉米基因组的主要组成部分,在调节基因组进化和适应方面发挥着关键作用。所有12个FIL显示出高度相似的转座元件(TEs)组成,占整个基因组~83.94-85.56%。在反转录转座子中,Gypsy(37.51-41.44%)和Copia(16.36-19.81%)超家族较丰富。Gypsy在着丝粒周围区域富集与基因分布呈负相关。而Copia在染色体臂富集,Helitron转座子的分布与Copia相似,并在着丝粒周围区域大量减少。Copia和Helitron与基因分布呈正相关。Helitron TEs位于接近基因的位置,尤其是外显子中,表明了Helitron-like转座子有助于基因复制、外显子重排,从而导致基因组进化和多样化。超过50%的DTT TEs和DTH TE位于基因附近(约2kb区域)。平均20.1%的DNA/DTH、23.1%的DNA/DTT TEs、16.8%的MITE/DTH TE和32.5%的MITE/DTT TE位于gene body中。平均4.7%的DNA/DTH、4.4%的DNA-DTT TEs、3.4%的MITE/DTH TE和3.1%的MITE/DTT TE在外显子区。平均3.4%的DNA/DTA和2.2%的MITEs/DTA TE位于外显子中,比DTH和DTT TE的频率低。这些结果表明,不同的TEs具有不同的分布和换位模式特征,它们的活动共同促成了玉米基因组的进化和多样化。
2、14个玉米骨干自交系泛基因组构建
通过14个玉米泛基因组构建,将在所有玉米中都检测到的基因被定义为核心基因家族(core ;18525个,42.57%),在12~13份玉米中检测到的被定义为软核心基因家族(softcore;9976个,22.92%),在2~11份中检测到的被定义为可变基因家族(dispensable;14992个,34.45%),只有一份中检测到的被定义为特有基因家族(private;25个,0.06%)。
通过模拟分析发现,总基因集随着基因组的增加而增加,当n=12时接近稳定(图2a),表明这14个玉米足够具有代表性。核心基因的百分比随着玉米数量的增加而下降,但即使当n=14时也没有稳定下来,表明玉米拥有大量的非共享基因。
随后,作者使用RNA-seq数据进一步研究了不同pan-gene的表达谱。总体表达丰度情况从核心、软核心到可变基因逐渐降低,表明核心基因具有稳定和更广泛的表达谱,而可变基因具有较低水平、组织特异性、品系特异性或条件依赖性表达。核心基因编码区中非同义变与总突变的比率低于软核心和可变基因(图2f)。可以注释的基因比例从核心、软核心到可变基因逐渐减少(图2g)。以上结果表明,核心基因在进化上更加保守。
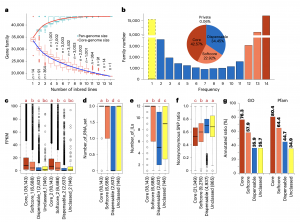
图2 14份玉米泛基因组分析
3、14个FILs基因组的共线性分析和遗传变异研究
接下来对14个FILs基因组进行成对的共线性分析和遗传变异研究,发现14个基因组之间平均共享56.31%的one-to-one共线性块。将13个FILs的所有one-to-one共线性块与B73_v4比对,发现这些FILs中存在广泛的遗传变异;并在保守区域发现了几个与玉米驯化或传播相关的基因,包括TGA1、TB1和ZmCCT10。此外,该分析也为杂种优势群的形成提供了参考。研究发现,亚热带FIL S37与13个温带FILs的共线性和IBS均低,温带FILs间的共线性较高(图3a)。值得注意的是,三种TSPT型FILs与其他FILs的共线性较低,说明其具有独特的基因组组成和特征(图3a)。揭示了TSPT类群与其他类群(特别是PA-和X-品系)可以形成优良的杂种优势模式的潜在机理。
对14个FILs基因组成对比对分析,在共线性区域,平均检测到8,489,166个SNPs、1,223,484个small InDels (< 50bp)和37,459个SVs(50bp-10kb)(图3b)。并且,本研究开发了一种方法用来鉴定13个FILs和B73之间的大片段SVs(大于100 kb),平均检测到337个大片段SVs,其中有12个的SVs大于1 Mb,包含先前报道的存在于B73和Mo17间的2.9 Mb的SV,含有玉米甘蔗花叶病毒(SCMV)抗性基因ZmTrxh。
另外,该研究也在14个FILs中鉴定了PAVs,平均鉴定到16,152个PAVs和184个PAV基因(图3b)。GO富集分析表明,PAV基因在防御反应、DNA整合、蛋白质磷酸化、萜类生物合成过程、激素介导的信号通路、细胞氧化解毒等生物过程中富集。将19个野生近缘种和23个地方种的重测序数据与14个FILs基因组进行比对,在至少一份大刍草或地方种质中,平均检测到93.4%的FIL特异性序列的密切相关同源物,与之前的报道一致,这些PAVs中的大多数都是来源于玉米祖先种中存在的变异。
14个FILs的共线性基因中,1,964个在基因内部(外显子和内含子)没有出现任何遗传变异,归类为结构保守基因。GO富集分析表明,保守基因富集的生物过程主要与植物的基本生命过程有关,如端粒组织或维持、DNA结构改变、重组、代谢与修复、染色体组织、细胞器组织等。值得注意的是,平均有7,390个共线性基因具有较大效应突变,如起始密码子或终止密码子突变、剪接供体或剪接受体突变、移码突变或过早终止密码子突变,表明这些FILs共线性基因之间的仍具有较大的遗传多样性。
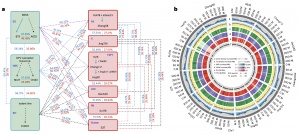
图3 14个FILs基因组的比较分析
4、SVs对不同杂种优势群的表型贡献
据报道,ZmCCT10、ZmCCT9、VGT1和ZCN8几个基因的自然变异是玉米从热带地区向温带地区传播的重要驱动力。350份温带ILs的PCR基因分型分析发现,早花型单倍型在ZmCCT9、VGT1和ZCN8上的累积效应与这些杂种优势群的开花表型一致。因此,ZmCCT9、VGT1和ZCN8对温带玉米杂种优势群的开花时间分化有一定的贡献,在未来的温带玉米育种中,这3个基因可作为提高PA、TSPT、PB品系开花期改良的位点。
玉米穗行数(KRN)是影响玉米产量的重要因素。先前的研究已经报道了位于UB3下游约60 kb的一个1.2 kb的转座子片段的插入,是一个主效的 QTL KRN4的重要功能变异。本研究发现SS 品系具有高KRN的有利单倍型,揭示1.2kb转座子插入可能是SS品系中更高KRN的重要原因。之前研究发现,一个编码丝氨酸苏氨酸类受体蛋白激酶的基因KNR6,可以调控玉米产量。本研究证实穗长(EL)和行粒数(KNR)与KNR6 5′-UTR区域一个Harbinger-like转座子插入引起的两种单倍型显著相关。350份温带ILs的PCR基因分型表明该SV可能是NSS和TSPT品系EL和KNR差异的主要原因。
利用Xu178(抗玉米粗缩病,MRDD)和HuangC的RILs群体进行玉米粗缩病(MRDD)抗性QTL定位,在8号染色体上检测到一个主效QTL。并且发现在14个FILs中,只有Xu178在抗性基因ZmGDIα中存在功能获得突变(由Helitron转座子插入引起)。进一步分析表明Helitron转座子是Xu178抗MRDD的重要原因。此外,分析表明Mo17和OH43的第6号染色体均含有约2.9Mb的大片段SV,可能是Mo17和OH43对SCMV易感性的基础。350个温带ILs的PCR基因分型进一步表明大片段SV是区分不同抗病性品系的主要位点。
5、eQTL研究确定基因表达变异和农艺性状相关的SVs
SVs通常影响基因的表达,本研究对350个温带ILs中的131个ILs的幼穗进行RNA-seq测序。通过eQTL分析,共鉴定出306,868个近端eQTL位点。ZmLOX3是引起玉米茎腐病和穗腐病的真菌病原菌镰孢菌(Fusarium verticillioides)的主要感病因子,是抗病性的负调控因子。作者选择一个与ZmLOX3相邻的eQTL进行表达分析,发现了两个SV变异(SV151和SV363),均与ZmLOX3的表达显著相关。基于SV151和SV363,鉴定出ZmLOX3的4种单倍型(Hap-0/0, Hap-0/363, Hap-151/0和Hap-151/363),Hap-0/363为有利单倍型,Hap-151/0为不利单倍型,为未来育种中利用这些SVs提高穗腐病抗性提供了潜在的新靶点。
eQTL分析还检测到两个近端eQTL,分别与Zm00001d006055和Zm00001d011140的表达水平相关 (图4a和5a)。Zm00001d006055的同源基因调控花的形态发生,Zm00001d011140的同源基因在调节植物发育中发挥重要作用。350个温带ILs的基因分型显示,Zm00001d006055(图4a-4c)的6 bp InDels和Zm00001d011140(图5a-5c)的92 bp SVs位于已报道的染色体开放区域(OCRs),与各自的表达变异显著相关。此外,田间试验表明,与野生型相比,功能缺失突变体Zm00001d006055(图4d-4f)和Zm00001d011140(图5d- 5f)较其野生型材料分别表现为雄穗分枝显著减少和穗位高显著降低的表型。上述结果表明,Zm00001d006055和Zm00001d011140分别对雄穗分枝数和穗位高有正向调控作用。等位基因频率分析显示,PA品系中含有6 bp InDel的单倍型的频率(21.1%)远高于TSPT品系(4.3%),NSS品系中含有92 bp的单倍型的频率(31.1%)远高于TSPT品系(4.3%)(图4g和5g),可能是这些亚群间对应性状分化的重要原因。
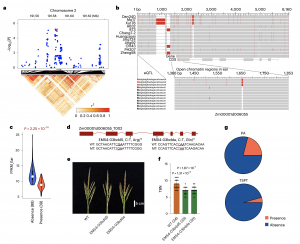
图4 Zm00001d006055的功能验证
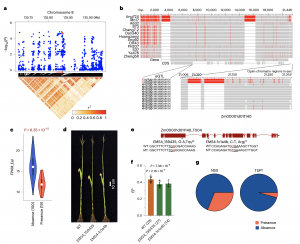
图5 Zm00001d011140的功能验证
6、杂种优势与基因组变异的相关性
为了研究玉米非核心基因组变异与杂种优势之间的关系,作者得到了14个FILs双列杂交的91个F1S,并收集了3个不同环境下的产量数据。表型分析表明,以单株产量(GYPP)计算,所有F1杂种均表现出超过双亲平均值的中亲优势(MPH)和超过较好亲本的超亲优势(BPH)(图5a)。
之前研究报道了杂种优势与分子遗传距离(GD)存在弱正相关关系。本研究为了研究产量杂种优势与遗传变异之间的关系,首先将基因组划分为共线性和和非共线性区域。相关分析显示,共线性区域长度与GYPP的BPH呈显著负相关(图5b)。共线性区域的SNPs和small InDels仅与GYPP的BPH呈弱相关 (图5c和5d)。相反,非共线性区域长度、SV_Number和PAV_Number均与GYPP的BPH呈显著正相关(图5e-g)。其中,SV_Number与GYPP杂种优势表现出强的相关性(图5f)。SVs可以进一步分为插入或缺失、串联收缩扩张和重复收缩扩张。其中,插入缺失与GYPP杂种优势的相关性强。综上所述,表明SVs的数量对GYPP的贡献很显著,支持了SVs在杂种优势中起主导作用这一观点。

图6 91个双列杂交F1S亲本杂种优势与不同类型遗传变异的相关性分析
7、ZmACO2和ZAR1中的SVs有助于杂种优势的形成
近期研究报道的乙烯生物合成关键基因ZmACO2,为穗长和籽粒产量主效QTL KNPR6的功能基因。前期研究对214个不同自交系的重测序分析和候选GWAS分析确定了5个与穗长显著相关的多态性变异,这些变异处于高度连锁不平衡状态。这五个多态性位点形成了两种不同的单倍型:Hap-SL17和Hap-Ye478。与携带Hap-SL17的自交系相比,携带Hap-Ye478的自交系穗长和行粒数更多,并且在携带Hap-Ye478的自交系中ZmACO2的表达显著低于Hap-SL17的自交系。
为了评价这些遗传变异对杂种优势的影响,作者对14个FILs进行了单倍型分析,除了Hap-SL17和Hap-Ye478之外,还得到一种新的单倍型(命名为Hap-Jing92)(图6a)。具有Hap-Ye478/Hap-SL17或Hap-Ye478/Hap-Jing92杂合单倍型的F1杂种的GYPP_MPH和GYPP_BPH的均值均显著高于各自纯合单倍型的成对亲本(MPP)的平均值(图6b-d)。这些结果支持ZmACO2可能以超显性方式促进杂种优势的产生。为了进一步验证ZmACO2在杂种优势中的作用,将两个近等基因系NIL-Ye478和NIL-SL17组合成F1杂交,并将其产量与两个亲本进行比较。结果表明,F1杂种的穗长、行粒数和产量均显著高于亲本NILs,验证了ZmACO2在调节杂种优势方面的超显性效应(图6e-6g)。
之前的一项研究报道,玉米ARGOS1(ZAR1)转基因过表达后可提高玉米生长量、子粒产量和抗旱性。分析表明ZAR1是一个杂种优势基因,同样可能以超显性方式促进杂种优势。为了进一步研究ZAR1单倍型对产量杂种优势的影响,对14个FILs中ZAR1的单倍型分析发现,除了先前报道的Hap NSS和Hap SS外,还有一种TSPT品系特有的第三种单倍型(将其命名为Hap TSPT)(图6h)。具有杂合单倍型(Hap-NSS/Hap-TSPT或Hap-SS/Hap-TSPT)的F1杂种的平均GYPP、GYPP_MPH、GYPP_BPH均显著高于各自纯合单倍型的亲本的平均值。但是具有Hap-NSS/Hap-SS杂合单倍型的F1杂种的GYPP、GYPP_MPH、GYPP_BPH的均值与其两个纯合单倍型的MPP均值没有显著差异(图6i-k)。这种不一致性可能是由于ZAR1单倍型的复杂环境互作造成的。总之,前期研究和本研究的结果强烈表明ZAR1以超显性方式促进杂种优势,其启动子和编码区域的自然变异有助于杂种优势的产生。
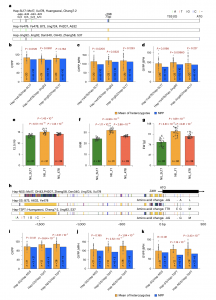
图7 ZmACO2和ZAR1的自然变异有助于杂种优势的形成
总 结
本次研究完成了12个核心玉米骨干自交系基因组组装并结合已公布B73、Mo17基因组构建了玉米泛基因组。通过基因组比对与变异检测并结合eQTL定位和关联研究,揭示了玉米基因组结构变异与杂种优势群的分化和性状调控密切相关。发现玉米杂种优势与父母本基因组的非线性区段及存在的结构变异呈显著正相关。鉴定了两个玉米杂种优势关键基因(ZAR1和ZmACO2),证明了Complementation和over-dominance都参与玉米杂种优势。本研究结果促进了对不同玉米杂种优势类群的基因组和表型分化的遗传基础的理解,为阐明玉米杂种优势机理迈出了坚实的一步,为玉米的全基因组选择育种研究提供了有力支撑。
相关阅读:
- 年度盘点|2022年百迈客基因组再创佳biomarker.com.cn/archives/28102绩!
- 林业果树研究所完成板栗泛基因组及数据库搭建
- 苦瓜T2T基因组发布!该研究揭示了‘金铃子’基因组组成和进化关系以及果实生长发育的分子调控机制
- 目前世界大规模牛泛基因组和基因组结构变异数据库构建完成!
- Natl Sci Rev:柑橘无融合生殖演化研究
- 【Nature Genetics】甘蔗细茎野生种种质资源演化及功能基因挖掘
- 动植物de novo测序的应用_15篇文献案例
- Commun Biol共同作者|橘小实蝇染色体水平基因组成功破译!入侵防控获得重大突破
- 2021百迈客denovo合作年度影响因子突破320+
- Hortic Res园艺TOP期刊-百迈客denovo合作将破20篇!植物基因组这么做!
- Mol Plant+Genome Biol共同作者|木薯如何靠基因组和群体半年破双十?
- IF=16.24 棉花转座子与基因组三维结构进化的奥秘
- 连续三篇,别掉队!现在流行全部denovo!
- 《Mol Plant》泡桐速生及泡桐丛枝病形成的机理研究-基因组精细图丛枝植原体完成图
- Nature Genetics高质量黑麦基因组揭示重要的农艺基因
- ADV SCI人工合成异源四倍体“金瓜”基因组揭示多倍化机制
- PNAS|金鱼的进化起源和驯化史
- CELL重磅|百迈客Hi-C助力榕-蜂共生体系基因组破译!
- Nature Plants|“小米”基因组破译–为C4禾谷类研究提供新型理想模式植物
- 高质量中国真蛸基因组带您走进paper新世界
- MER | Nanopore完成超高质量海洋鱼类基因组
- NC|雷山髭蟾基因组揭示两栖动物性二态分子机制
- 开心果基因组,见刊GENOME BIOLOGY!
- 《ISCIENCE》喜马拉雅旱獭基因组揭示其缺氧和冷适应机制【含文献原文】
- 《NATURE GENETICS》三代异源四倍体陆地棉和海岛棉基因组破译
- 《NATURE GENETICS》 百迈客助力破译同源四倍体甘蔗基因组
- 河南农大殷冬梅教授团队首次发布异源四倍体花生基因组
- 百迈客亚洲棉科研成果在NATURE GENETICS上发表




 京公网安备 11011302003368号
京公网安备 11011302003368号