
百迈客单细胞测序
单细胞转录组测序(scRNA-seq)是在单细胞水平进行高通量基因表达谱检测,对复杂细胞群深入分析,表征单个细胞的表达谱,避免单个细胞的异质性生物学信息被大量细胞的均质化覆盖。
导读
用人多能干细胞(hPSC)形成种间嵌合体已成为在体内评估hPSC多能性的有效方案,并且可能在再生医学,包括移植器官和组织再生中发挥重要作用。使用小鼠和猪胚胎的研究表明,hPSCs并不能有力地促进与人类进化距离较远的物种的嵌合体形成。本研究主要是在体外培养的食蟹猴(Macaca fascicularis)胚胎中研究人扩展多能干细胞(hEPSC)的嵌合能力,证明hEPSCs能够在食蟹猴胚胎中存活、增殖,并绘制植入前后细胞图谱。同时还发现了种间细胞相互作用,这些事件可能有助于塑造嵌合胚胎内人类和食蟹猴细胞的独特发育轨迹。本研究结果可能有助于更好地了解早期人类胚胎发育和灵长类动物进化,并制定策略以改善进化距离较远的物种的人类嵌合体的形成。
实验方法
材料:注入tdTomato+ hEPS细胞的食蟹猴早期囊胚
方法:单细胞转录组测序
研究结果
1、体外人猴嵌合囊胚的形成
为确定非人灵长类动物中hPSC的嵌合能力,本研究使用了通过细胞重编程产生的特征明确的hEPSC系iPS1-EPSC,它在小鼠胚胎第10.5天(E10.5)表现出优于其他hPSC的嵌合性。与之前的报告一致,iPS1-EPSCs表现出圆顶形集落形态,并表达核心多能性转录因子OCT4、NANOG和SOX2(图 S1A)。为了生成人猴嵌合胚胎,对食蟹猴的早期囊胚(受精后 6 天 [d.p.f.6])注射了25个用tdTomato(TD)标记的iPS1-EPSC,注射的胚胎培养到晚期囊胚阶段(d.p.f.7)进行分析,发现食蟹猴囊胚内hEPSC的增殖很明显。总的来说,在所有d.p.f.7的食蟹猴囊胚中检测到TD+ iPS1-EPSCs(100%,n = 132)(图 S1B)。
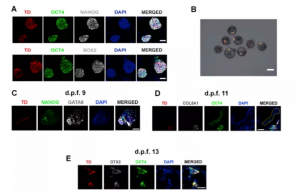
图S1 宿主食蟹猴胚胎中hEPSCs的谱系规范
2、人猴嵌合胚胎的转录图谱
为进一步描绘人猴嵌合胚胎的发育轨迹,进行了scRNA-seq以分析不同发育阶段的人和猴细胞的转录组。胚胎分离后,使用荧光显微镜手动收集单个人类(TD+)和猴(TD-)细胞并进行scRNA-seq。对离体培养过程中不同时间点从嵌合胚胎中分离的227个人的和302个食蟹猴的细胞进行了测序(d.p.f.9-d.p.f.17)。TD表达和比对到人类或食蟹猴基因组的reads用于进一步确认每个细胞的来源物种(图 S2A 和 S2B)。严格过滤后,200 个人的和 272 个食蟹猴的细胞用于进一步分析。每个细胞平均检测到 9,798 个基因(每百万转录本【TPM】>0)和27,936,953条reads,在人和猴细胞之间检测到的基因数量和reads没有统计学差异(图 S2C)。为进行比较,本分析中还选择了已发表的食蟹猴和人类胚胎细胞的scRNA-seq数据集。为避免不同数据集的批次影响,本研究使用“锚定”方法去除批次效应(图 S2C)。
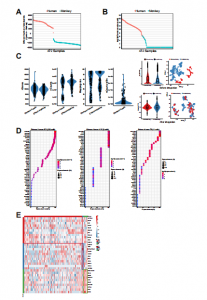
图S2 细胞物种来源鉴定和QC
对scRNA-seq数据进行了t-SNE分析,确定了所有样本(嵌合胚胎和对照胚胎)中存在4个主要细胞cluster:EPI、HYP、TE和EXMC(图 1A、1B和S2D)。通过细胞特异性marker分析,发现人和食蟹猴之间存在保守性(图 S2E)。嵌合胚胎中这些细胞类型的存在表明宿主胚胎的发育基本上不受注射的hEPSCs的影响。系统发育树分析(基于基因表达水平)显示,虽然嵌合胚胎中的大多数食蟹猴细胞分离成不同的细胞类型特异性簇(EPI、HYP和TE),但嵌合人类HYP-和TE-like 细胞与 EPI-like 细胞聚集在一起(图 1C)。因此,嵌合食蟹猴细胞比引入的hEPSC表现出更强的谱系分离。与IF结果一致,在scRNA-seq数据(图 1A 和 1D)中鉴定到的人类TE-like细胞很少,因此被排除在后续分析之外。这些结果表明,hEPSCs在被引入食蟹猴早期囊胚并进行离体胚胎培养后,可以分化为几种植入前和植入后早期细胞类型。
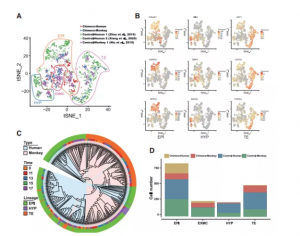
图1 人猴嵌合胚胎的单细胞转录图谱
3、hEPSCs在人猴嵌合体发育过程中的转录组动力学
首先基于所有细胞的转录组学特性构建了一个导向图(SPRING),所有细胞分为三个分支:EPI、HYP和TE(图 2A),明确了嵌合体和对照(人和食蟹猴)胚胎之间基因表达模式的相关性(图 2B)。当嵌合人类细胞与对照人类(0.460)或对照食蟹猴(0.459)细胞进行比较时,获得了类似的相关系数(图 2B,右图),但与对照胚胎相比,嵌合猴细胞的相关系数高于嵌合人类细胞(图 2B,左图);接下来发现嵌合人EPI-like细胞与人胚胎中的EPI细胞相似,而嵌合人HYP和EXMC-like细胞分别与嵌合猴HYP和EXMCs细胞相关系数最高(图 2C)。同时发现嵌合人类EPI-like细胞逐渐倾向于嵌合猴EPI细胞,R2值从0.363(植入前EPI [Pre_EPI])增加到0.464(植入后EPI [PostL_EPI]),再增加到0.693(原肠[Gast]细胞)(图 2C)。以上结果表明猴胚胎微环境对人类细胞的基因转录状态产生影响,反之亦然。
由于食蟹猴的细胞在人类细胞存在下表现出转录组的变化,接下来分析了嵌合胚胎中食蟹猴细胞的发育动态。首先确定了嵌合猴胚胎和对照猴胚胎之间的差异表达基因(DEG)。EPI细胞、HYP细胞和EXMCs细胞与对照胚胎相比,嵌合体细胞中分别有424、7和241个基因下调,5、2和13个基因上调(图 2D和S3A)。GO和KEGG富集分析确定了在嵌合猴EPI细胞、HYP细胞和EXMCs细胞中上调和下调基因的信号通路,如Hippo和转化生长因子β(TGF-β)信号通路分别在嵌合猴EPI细胞和EXMCs细胞中下调(图 2E)。
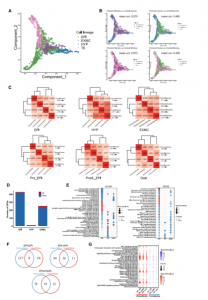
图2 嵌合胚胎的发育轨迹
已经证明食蟹猴EPI细胞的转录组谱在嵌合胚胎中发生了改变,接下来研究了食蟹猴EPI胚胎生态位是否也受到人类细胞的影响。CellPhoneDB(v2.0.1)可鉴定嵌合胚胎和对照胚胎中EPI和其他谱系(HYP 和 EXMCs)之间细胞的潜在相互作用。结果发现与对照胚胎相比,嵌合胚胎中有更多的配体-受体相互作用(如,在猴EPI细胞中检测到117个 [嵌合] 与10个 [对照] 特异性配体-受体相互作用)(图 2F 和 S3B)。进行KEGG分析发现,嵌合胚胎中被加强的信号通路包括磷脂酰肌醇 3-激酶 [PI3K]-Akt、丝裂原活化蛋白激酶 [MAPK] 信号通路和WNT信号通路(图 2G)。使用相同的方法,还确定了嵌合胚胎内人和猴细胞-细胞的相互作用,如FGF5-FGFR4、NOTCH4-JAG2、WNT2B-FZD4、WISP3-SORL1和PLXNB2- PTN(图 S3C 和 S3D)。结果表明,嵌合胚胎内的细胞间相互作用得到加强,并可能导致其他信号通路的激活。
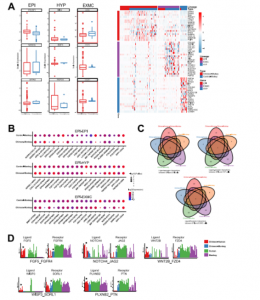
图S3 嵌合胚胎与宿主胚胎食蟹猴细胞对比分析
4、嵌合的人类EPI-like细胞显示出独特的发育轨迹
EPI发展的特点是进行多能性转变,可能在物种之间表现出不同的动态。适当的EPI分化对于嵌合体的形成和发育至关重要,本研究对嵌合胚胎内人类EPI-like细胞的谱系分化进行了研究,并将其与体外培养的人类和食蟹猴胚胎的数据集进行了比较,结果人类EPI-like细胞在植入前、植入后和原肠胚形成阶段被鉴定,并且在每个阶段表达不同的标记物(图 3A),桑基图也显示了相同的人类EPI-like细胞发育轨迹。接下来观察到hEPSCs更类似于早期PostE_EPI和PostL_EPI细胞。嵌合体中人PostL_EPI-like细胞与制备的PSC的相关性高于naive PSC。为了进一步研究 hEPSC、嵌合体人类 EPI-like细胞和宿主猴EPI细胞的转录动力学,本研究进行了RNA速度和Slingshot分析(图 3B),结果发现两种不同的RNA速度向量模式:嵌合体人类PostL_EPI-like细胞向量较长,而原肠胚细胞向量较短;宿主猴PostL_EPI细胞缺乏长向量,而原肠细胞具有长向量(图 3B,左边两张图)。这些结果表明嵌合人类EPI-like细胞的发育延迟。Slingshot分析显示,hEPSCs在注入食蟹猴囊胚后,是从EPSCs到PostL_EPI再到原肠胚的发育轨迹(图 3B,右图)。为了进一步描绘嵌合人类EPI-like细胞的发育轨迹,将所有与EPI相关的人类和猴子reads映射到一个共有基因组,并使用先前报道的方法预测物种之间EPI的发育轨迹,与RNA速度分析一致,发现嵌合人类EPI-like细胞比来自宿主猴、对照猴和人类胚胎的EPI细胞分化得更慢(图 3C)。这些结果表明,hEPSCs向EPI谱系的特化和/或分化效率低于胚胎细胞。
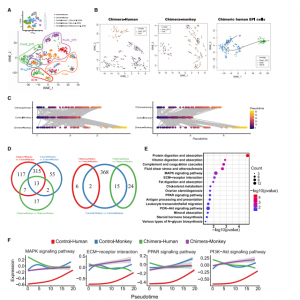
图3 食蟹猴胚胎中嵌合人EPI-like细胞的发育轨迹




 京公网安备 11011302003368号
京公网安备 11011302003368号