

2021年4月,中国热带农业科学院李开绵研究员团队牵头完成了高质量单倍型木薯基因组,相关研究成果“Allele-defined Genome Reveals Biallelic Differentiation during Cassava Evolution”发表于国际期刊Molecular Plant(IF:13.162)。近日,该团队再次牵头完成了木薯群体研究,该研究以“Resequencing of 388 cassava accessions identifies valuable loci and selection for variation in heterozygosity”为题见刊Genome Biology(IF:13.584)。百迈客有幸作为共同作者参与这两项研究。接下来,随小编一起来看看两篇文章如何克服物种高杂合的困难,并以此为突破点开展系列研究。
木薯基因组

文章亮点
本研究首先通过PacBio平台构建具有“冗余”特征的基因组,再进一步结合Hi-C互作信号,将二倍体两套的基因组进行拆分,最终获得具有18对同源染色体的双单倍型基因组。前期研究结合本次同源对基因组的差异比较,进一步说明了同源染色体之间不同遗传位点的组合对生物表型有重要影响。该研究为广大科研工作者提供了一个重要的研究思路:无双亲本背景下,构建高杂合物种的单倍型基因组。通过单倍型的分析,为解析物种重要性状形成的遗传机制奠定了重要基础。
研究背景
通过种间或种内杂交产生杂合子基因组有助于维持植物多样性并成为新物种的潜在来源。由于缺乏高质量的参考基因组和有限的基因组资源,木薯基因组学和育种研究相对滞后。木薯(Manihot esculenta Crantz;2n=36)是非洲和其他热带地区的主要作物,其具有很高的杂合性,是研究双等位基因分化的理想系统。
材料方法
基因组:木薯栽培品种SC205;~98.5X PacBio;~35X Hi-C
转录组:种下后100-340天(每隔40天取样)的块根;以及根茎叶和块根组织
研究结果
木薯基因组组装
作者通过PacBio和Hi-C技术进行基因组的初始组装,组装大小为1.1Gb(流式预估单套大小770.3 Mb),表明在基因组的杂合区域组装了两个单倍型。作者通过自对z确定了冗余区域,并将非冗余木薯基因组作为参考,称为SC205-ref。组装基因组的连续性指标ContigN50=1.1 Mb,Scaffold N50=34.5 Mb,并经过BUSCO(92.8%)评估、LAI评估(18.46)、高密度遗传图谱(相关系数ρ>0.99)等多种评估方式,表明了本次构建了一个参考级的高质量基因组。
进一步使用ALLHiC算法进行等位基因定义的基因组组装,组装1.52Gb木薯基因组,构建了18对同源染色体高质量单体型基因组图谱,鉴定了24128个双等位基因(Bialleles)。并通过Hi-C信号、同源染色体间共线性比对、遗传图谱比对、二代/三代Reads回比、同源对BUSCO评估、双等位基因鉴定等方式对本次单倍型基因组进行验证。
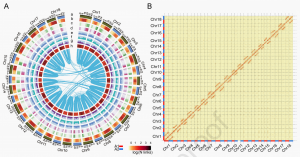 图1 木薯双单倍型基因组特征
图1 木薯双单倍型基因组特征
不对称演化
通过同源染色体比对,鉴定到3,107,171 个SNPs、 500,018 个indels、57,643 个PAVs以及632 个倒位。高杂合度区域的进化速度比其他区域更快。
蛋白质编码基因、不同等位基因和显性表达的双等位基因的密度,发现它们都倾向于富集在高度杂合的区域,通常位于染色体的一端或两端附近。通过对不同种植时间和组织部位的转录组分析发现除了第1、15和17对染色体外,几乎所有同源染色体对中,双等位基因间广泛发生表达不平衡,并且淀粉和蔗糖代谢途径上的双等位基因表现出显著的表达分化;
等位基因进化
显性表达的等位基因的启动子和CDS的序列差异显著大于同等表达的双等位基因。根据非同义和同义核苷酸替换的比率(Ka/Ks)将等位基因对分为7个亚群。等位基因组进化分析揭示了基因组快速进化可能是木薯高杂合性形成的重要驱动力,且基因组方向性选择驱动等位基因表达分化。
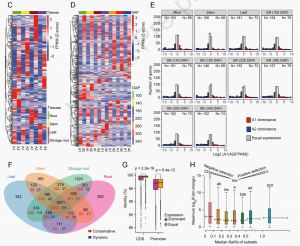
图2 木薯等位基因组表达与进化分析
总结
本研究通过构建高质量的木薯单倍型参考基因组,深化了对木薯双等位基因变异遗传基础的理解,有助于探索双等位基因的分化和表达优势及其潜在的进化驱动力,为木薯改良的战略性开发提供了可能。这些基因组资源将为创新木薯和其他高杂合作物的育种策略提供新的见解。
木薯群体进化

文章亮点
本研究突破前期纯合子等位基因变异与物种关键农艺性状的相关性研究,进一步聚焦高杂合物种的等位基因变异。发现不同基因位点的纯合与杂合变异涉及到块根重量、淀粉含量、抗病性等多种木薯关键的农艺性状。阐明了杂合位点的
保持对于优异性状的形成具有重要贡献,为高杂合物种研究提供了一个新颖的研究思路。
研究背景
许多野生物种通过人工选择驯化为栽培作物,随着人类迁移、定向选择和进一步的性状改良以满足人类需求,栽培作物在性状上获得了区域特异性差异。利用基因组测序技术,我们可以追踪作物驯化和育种历史,从而更好地了解人类选择如何形成作物基因组。群体中基因型和表型之间的关联显示纯合子等位基因变异与许多作物的关键农艺性状显著相关,从而加速了育种过程。然而,许多作物是高度杂合的,杂合性变异对农艺性状的影响尚不清楚。
栽培木薯是6000多年前从亚马逊流域的野生祖先Manihot esculenta ssp. flabellifolia驯化而来。木薯的驯化产生了包括年生长周期、高初始生长率、产量增加和高淀粉含量在内的特征,这使得木薯成为人类消费和工业生物燃料应用的理想能源。但由于基因组资源有限,木薯基因组学和育种研究相对滞后。群体基因组资源的开发将加速这一重要作物的遗传改良进程。
材料方法
代表来自全球15个国家主要木薯生产区的388份木薯种质(14份野生、38份地方品种和336份栽培品种);51份材料来自发表重测序数据,337份在本研究中测序。总共产生3.35 Tb数据,平均测序深度为9.45×。对本次测序的337份木薯,进行三年的33个形态性状GWAS分析。
研究结果
基因组变异图谱与系统发育关系
通过与参考基因组比对,共鉴定1,344,463个SNPs以及1,018,832个InDels。系统发育和主成分分析(PCA)将388份木薯材料分为两大类(图1c,d),第一组包括13个野生祖先,第二组包含所有地方品种、栽培品种以及自巴西的野生祖先FLA433-2(具类似木薯的块根)。结果支持了FLA433-2是木薯野生祖先种的假说。系统进化及FST、p的分析结果与考古证据相符,表明南美和非洲木薯种群之间以及非洲和亚洲种群之间存在着密切的系统发育关系,提出了木薯从南美洲到非洲再到亚洲的传播驯化路径。
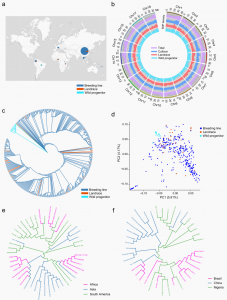
图1 木薯全基因组变异图谱及其亲缘关系
全基因组关联分析(GWAS)
GWAS分析鉴定出与23个重要农艺性状关联的52遗传标记。表皮类型(光滑或粗糙)和疤痕严重影响木薯块根的外观质量。10号染色体上Sc10g01040中的三个和Sc10g02050中的七个非同义SNP与块根表皮型相关。在10号染色体Sc10g01040、Sc10g01050以及3号染色体Sc03g001750、13号染色体Sc13g000920中携带杂合等位基因和杂合单倍型的木薯材料比携带纯合等位基因和纯合单倍型的木薯材料具有更高的块根平滑表皮频率。木薯地上部分的重量反映了植物的生长活力,并成为其用作饲料的一个重要因素;块根支链淀粉含量决定木薯的工业价值。5号染色体上Sc05g013530、11号染色体上Sc11g000910携带杂合子等位基因的材料表现出显著更高的地上重量和支链淀粉含量。2号染色体Sc02g008280携带AT或TT等位基因的材料具更高红色内皮颜色频率,18号染色体Sc18g013220携带杂合子AC等位基因的材料比携带纯合子CC或AA等位基因的材料显示出显著更低的螨感染指数。对这些候选基因的表达水平进行检测,发现都在块根发育的不同组织或阶段大量表达。种种结果表明,杂合性等位基因变异在关键农艺性状中的潜在作用,其中每个候选基因中携带杂合等位基因的木薯材料比携带相应纯合等位基因的木薯材料具有更理想的表型。
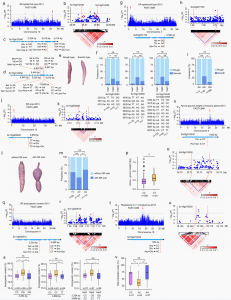
图2 GWAS鉴定与关键农艺性状相关的杂合性变异候选基因
作者鉴定了79个与块根中淀粉积累相关的高度杂合子片段,其中20个杂合块与6个GWAS信号重叠。此外,在1号染色体上携带高度杂合子片段的木薯材料比携带相应纯合子片段的木薯材料表现出更高的块根红色内皮细胞频率。这些结果进一步支持杂合性变异与关键农艺性状相关的假设。
木薯杂合性的选择特征
为了追踪杂合性的可能进化场景,作者根据参考基因组的杂合SNP将木薯基因组分为杂合区和纯合区。然后比较了374个品种的杂合区和纯合区的基因组多样性。在大多数(18条染色体中的16条)染色体中,杂合区具有显著高于纯合区的遗传多样性(π值)和正Tajima’s D值,表明平衡选择有助于维持基因组杂合性。在品种和野生祖先之间的大多数染色体(18个染色体中的14个)中,杂合区的FST值低于纯合区的FST值也支持了这一发现。检测到81个杂合度和核苷酸多样性降低的选择性位点,包含548个基因,这些基因在多种生物过程中富集,包括生长、发育、激素代谢和反应以及免疫相关过程。
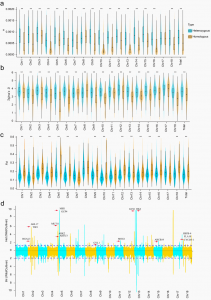 图3 木薯杂合性的选择特征
图3 木薯杂合性的选择特征
驯化使木薯块根的产量和淀粉含量显著增加。栽培品种的块根重和淀粉含量分别比野生后代高约60倍和3倍(图4a,b)。选择性清除分析结合转录组分析发现2号染色体MeT
R1周围杂合性降低的选择信号,并且MeTIR1在携带GG等位基因的材料中的表达显著高于携带CG和CC等位基因的材料(图4i),说明MeTIR1纯合变异促使块根淀粉含量提升。并通过瞬时过表达和沉默技术进一步验证了MeTIR1在木薯中的功能。进一步结合关联分析发现,对2号染色体携带MeAHL17 AA等位基因的材料,经选择驯化促使块根产量提升的同时导致木薯细菌性枯萎病抗性丢失。
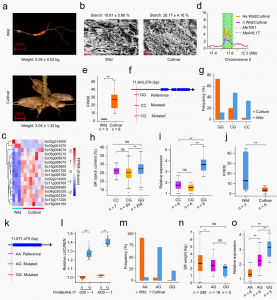
图4 木薯高淀粉含量块根驯化相关杂合性降低的选择作用
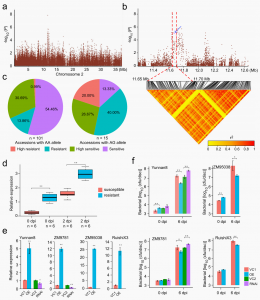 图5 木薯细菌性枯萎病候选基因MeAHL17的鉴定
图5 木薯细菌性枯萎病候选基因MeAHL17的鉴定
MeAHL17和MeTIR1位于2号染色体上相同的选择性清除区内。连锁不平衡分析发现,两个基因紧密连锁,携带AG(MeAHL17)和GG(MeTIR1)等位基因组合的品种显示出较高的淀粉含量和对木薯细菌性枯萎病的高抗性频率。
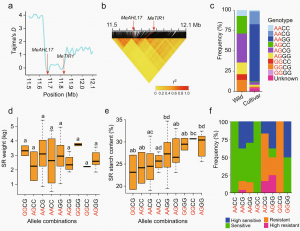
图6 MeAHL17和MeTIR1等位基因组合对性状变异的影响
总结
本研究提供了388份木薯材料的变异图谱,鉴定了23个农艺性状的52个位点,揭示了与关键农艺性状和木薯驯化相关的杂合性等位基因变异。检测到81个杂合度和核苷酸多样性降低的选择性位点,相关基因在多种生物过程中富集,包括生长、发育、激素代谢和反应以及免疫相关过程。人工选择MeTIR1和MeAHL17中的纯合等位基因有助于大淀粉块根的驯化。而在MeAHL17中选择纯合子等位基因与增加块根重和木薯细菌性枯萎病易感性相关。本研究将有助于阐明与关键农艺性状和木薯驯化相关的杂合性变异的遗传基础,并对木薯驯化过程中杂合性的变异提供了见解,为木薯和其他高杂合物种育种改良的战略发展提供依据。
写在最后:
大多数二倍体基因组组装都忽略了同源染色体之间的差异,将基因组组装成一个假的单倍体序列(嵌合型)。但随着研究的深入,发现仅单套的基因组数据难以完全演示该物种的全面信息,例如在木薯、马铃薯、茶树等物种中已有相关研究(Zhou Q et al., Nature Genetics.2020;Zhang X et al., Nature Genetics.2021; Wang P et al.,Horticulture Research)。同源多倍体中问题尤其明显,同源染色体之间不同遗传位点的组合对生物表型有重要影响,如动植物中的杂种优势、某些物种杂交不育现象等。
单倍型等位基因间差异对基因表达、功能及其表型都有着重要影响,大多数杂交品种的优势表型都受等位基因调控。通过单倍型的分析,可为解析物种重要性状形成的遗传机制奠定重要基础。
无论是单倍型基因组的构建还是泛基因组的构建,单个的样本往往不具有性状分析的普适性,需要进一步通过群体规模的大样本检测,挖掘与自然人工驯化、性状功能关联的关键位点,为物种的育种改良与相关基因研究奠定重要理论基础。
百迈客自2009年成立以来深耕于群体遗传研究,同时具有近10年基因组分析经验。现已在基因组、Hi-C、遗传图谱等技术上拥有实验+生信分析双保障。并与国内外70余所科研单位在国际期刊合作发表500余篇高质量文章,累计影响因子3000+。

为了答谢大家长期以来的信任与支持,百迈客特在年末之际,推出多项优惠活动,助力科研! 详情请咨询当地销售经理或拨打电话:400-600-3186




 京公网安备 11011302003368号
京公网安备 11011302003368号