



研究背景
黄瓜(Cucumis sativus L.)(2n=2x=14)是世界上重要的蔬菜作物,在驯化过程中经历了进化瓶颈,因此其遗传基础尤其狭窄。陈劲枫教授团队将栽培黄瓜(Cucumis sativus L., 2n=14)与野生酸黄瓜(C. hystrix Chakr., 2n=24)进行种间杂交,通过胚胎拯救和染色体加倍,甜瓜属人工异源四倍体新物种“金瓜”并命名为Cucumis hytivus Chen and Kirkbr.。金瓜具有明显的生长力旺盛、营养价值高等优势,同时还具有很强的环境适应性。
天然多倍体的亲本基因组通常是未知的,或是在多倍体形成后已经各自进化,因而无法系统地区分由种间杂交和基因组加倍所引起的变化。因此,对具有明确遗传背景的人工异源四倍体C.×hytivus的研究,可以解析多倍体化进程中不同过程的潜在机制,从而揭示异源多倍体在自然界和农业生产中广泛存在的深层含义。
材料方法
- 材料:C.×hytivus自交第14代(S14);测序:~69X PacBio+~104X Hi-C+Bionano;转录组辅助基因预测:Illumina RNA-Seq 及 PacBio Iso-Seq
- 材料:F1、S0、S4-S13 (组装用于比较基因组分析)测序:~80X Illumina X-TEN (scaffold N50=134~226 kb;contig N50 =47~73 kb)
研究内容
- 异源四倍体C.×hytivus(S14)的组装与注释
- 亚基因组优势分析
- 种间杂交、基因组加倍和二倍体化
- 多倍体化拓宽物种遗传基础,增强物种适应能力
主要研究结果
作者通过PacBio+BioNano+Hi-C+Illumina共组装获得530.8 Mb基因组(≈67%流式评估大小;≈77% survey评估大小;比对发现未装出的为重复序列),contig N50=6.5 Mb,并将组装的基因组序列挂载至19条假染色体上(挂载率97.23%;定序与定向率93.33%)(表1)。通过与二倍体C.sativus (CC)和C.hystrix(HH)染色体的C01–C07和H01–H12对应,将C.×hytivus染色体命名为Chc01–Chc07和Chh01–Chh12(图1)。
与已测序的栽培黄瓜基因组一样,金瓜中大部分的TE为长末端重复序列。C.×hytivus(S14)的Chc亚基因组(SubC)含有的TEs少于二倍体C.sativus(CC)基因组,而Chh亚基因组(SubH)保持了几乎相同的不同类型TEs的含量和比例。共预测了45687个基因,平均基因长3846bp,每个基因有5.26个外显子。BUSCO评估结果为90.90%,CEGMA评估结果为97.82%。在CC和HH二倍体基因组中鉴定的18882个同源基因家族中,在C.×hytivus(S14)异源四倍体中也鉴定了18428个(97.60%)(图2)。
表1 金瓜基因组组装结果
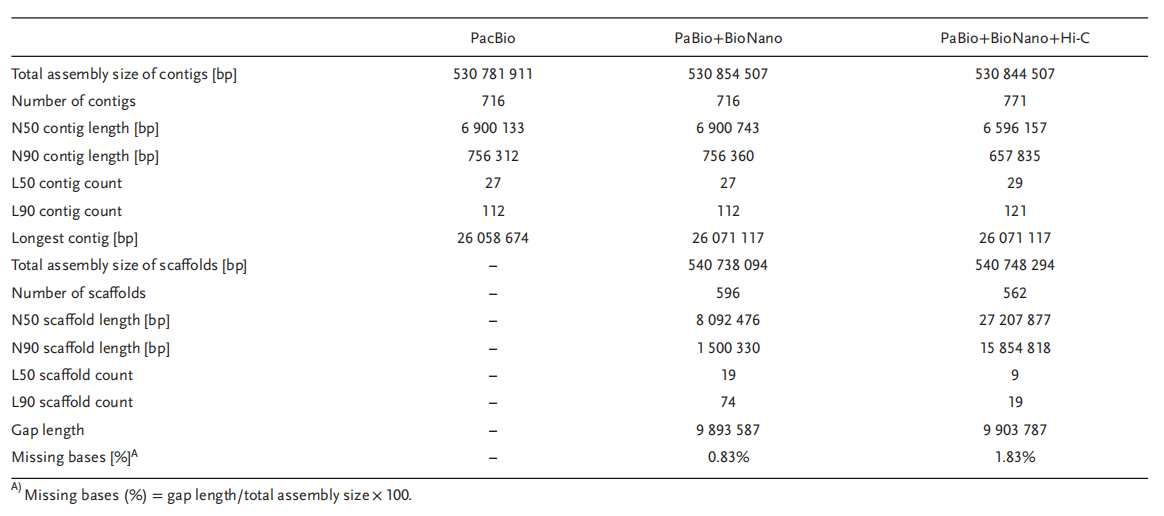
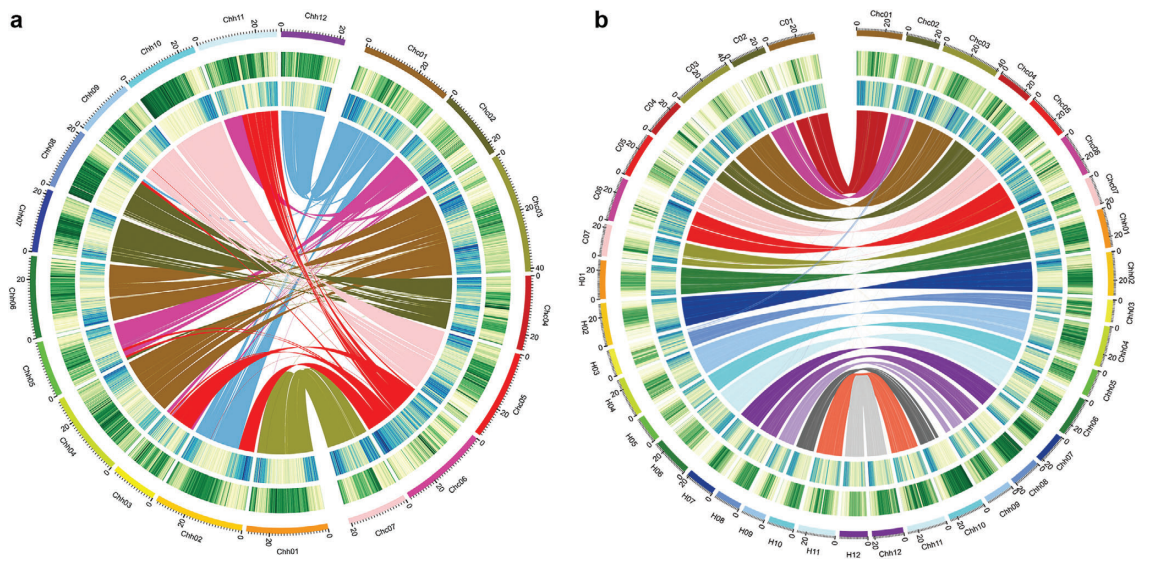
图1 金瓜亚基因组间共线性(左)及亚基因组与二倍体祖先共线性(右)
图2 异源四倍体后基因的变化
作者将C.×hytivus(S14)基因组分为Chc(203.36 Mb)和Chh(287.37 Mb)两个亚基因组,这两个亚基因组均小于相应的CC基因组(226.21 Mb)和HH基因组(297.49 Mb),分别包含的基因也比相应的亲本种少。与CC相比,在Chh亚基因组中检测到的SVs更多,并且基因丢失更多。通过序列深度分析及同源交换(HE)分析表明,在C.×hytivus(S14)基因组中分别丢失11个CC和146个HH基因,同时更多的HH序列被CC序列替换。共线性基因对在C.×hytivus(S14)亚基因组中的表达结果表明,CC显性基因的数量明显多于HH显性基因(图2C)。这些结果表明Chc亚基因组可能比Chh亚基因组更占优势。CC和Chc亚基因组的TE含量低于HH和Chh亚基因组,支持了由祖先细胞基因组中TE的差异密度驱动异源多倍体中亚基因组偏向分化的假说(TEs的失活扩散到附近的基因,TEs密度大的同源基因组的整体表达较弱,导致基因失活和最终丢失的可能性更大)。
异源多倍体化过程中的三个重要阶段(种间杂交、基因组加倍和二倍体化)对异源多倍体基因组的作用各不相同。
为了区分其效果,作者分别在种间杂种F1、金瓜初世代S0和其他早期世代(S4~S13)中对S14世代中预测的157个确认缺失的基因进行鉴定。结果表明,F1中有102个基因缺失,表明种间杂交事件发生后,核基因组序列立即被消除。而F1中有个别缺失的基因在S0中再次出现,表明基因组加倍在异源多倍体中的作用与种间杂交截然不同。比较DNA序列分析显示,相对于HH的叶绿体基因组,F1、S0和S4~S13的叶绿体基因组中共有195个SNPs和100个IndDels(图4)。其中绝大多数SNPs(73.85%)和IndDels(73%)在F1中检测到,推测多倍体中被广泛发现的亚基因组优势现象在种间杂交过程中就已经基本确定了,并在后续的进化过程逐渐累积。而在二倍体化过程中,基因组的变化也主要发生在最初的几个世代(S0~S3),之后的世代中仅观察到零星的核序列丢失(基因丢失),并且叶绿体基因组中没有发现新的SNPs或InDels。
六代植物(S4、S6、S8、S10、S12和S14)的减数分裂染色体行为显示,Chc亚基因组的单价体数量和滞后频率显著低于Chh亚基因组。表明该异源四倍体的Chh亚基因组存在一定的不稳定性,从而表现出较高的序列丢失率。Chh基因的不稳定性可能是导致C.×hytivus育性下降的主要原因。花粉可染性随着世代的增加而稳步增加,表明育性地逐步恢复(图3a)。
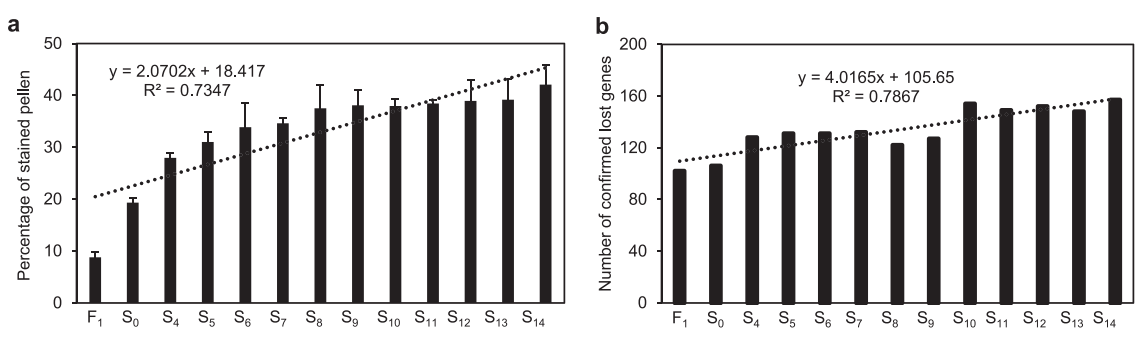
图3F1及早期世代花粉可染性研究(左)及缺失基因数(右)
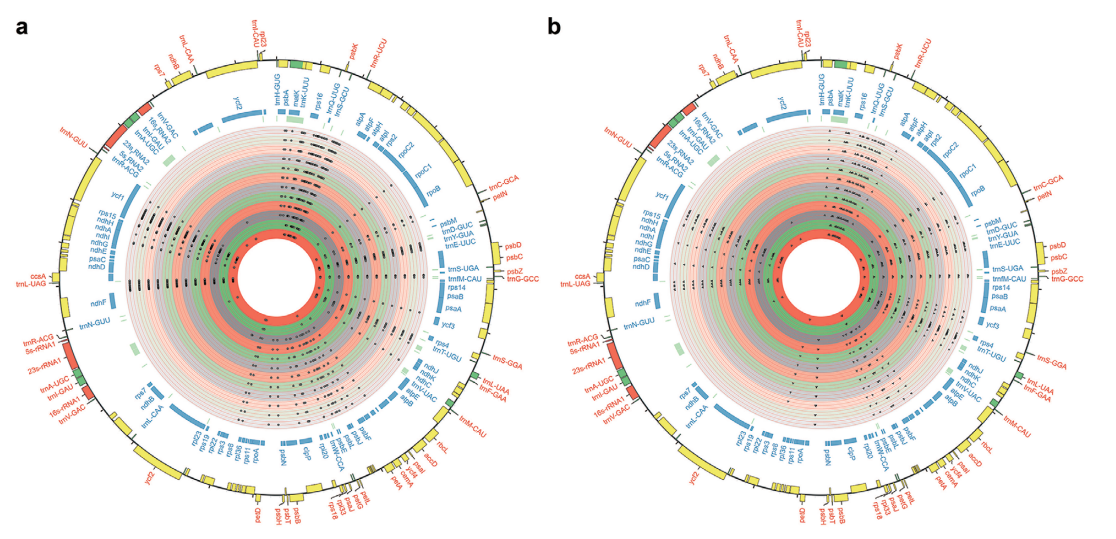
图4 F1及早期世代叶绿体基因组SNPs(左)和InDels(右)分布
人工合成的多倍体新物种金瓜不仅表现出明显的杂种优势(生长力旺盛、营养价值高),同时还具有很强的环境适应性,并且C.×hytivus(S14)对根结线虫(Meloidogyne spp.)的抗性与HH相当,且高于CC。于是作者对其进行了基因序列与表达量分析,发现金瓜中保留了来自双亲的大多数重复的NBS-LRR编码基因(携带核苷酸结合位点(NBS)和亮氨酸富集重复(LRR)结构的NBS-LRR类抗性基因),使其具有更强的抗病能力,比亲本物种具有更强的生存力。高温和对照条件下,C.×hytivus与其两个二倍体亲本叶片的生长、生理反应和转录组表达水平研究表明,主要参与光合生物的固碳、碳代谢、乙醛酸和二羧酸代谢相关的基因在C.×hytivus中显著表达,而亲本中则没有。证实了在热胁迫下,C.×hytivus叶绿素积累和光合作用的增强,进一步说明了金瓜具有耐热性。




 京公网安备 11011302003368号
京公网安备 11011302003368号