
2020年11月03日,北京百迈客生物科技有限公司合作项目金鱼基因组及群体驯化研究成果以“The evolutionary origin and domestication history of goldfish(Carassius auratus)”为题在国际知名期刊PNAS上在线发表。文中组装获得了高质量的金鱼基因组 (2n=100)的同时,通过比较基因组学研究解析了古代杂交事件产生了两个亚基因组;进一步通过185条代表性金鱼变种和16条野生鲫鱼的重测序分析,揭示了金鱼的起源,并鉴定了与驯化相关的受选择基因区域。研究中全面收集了金鱼品种,获得了与众多解剖特征相关遗传变异信息,其中包括区分传统金鱼进化的特征。除此之外,鉴定了酪氨酸蛋白激酶受体作为透明突变体的候选基因。为金鱼的基因组和多样性数据提供了丰富的资源,使得金鱼成为功能基因组学和驯化研究具前景的模型。
研究结果1:金鱼基因组组装
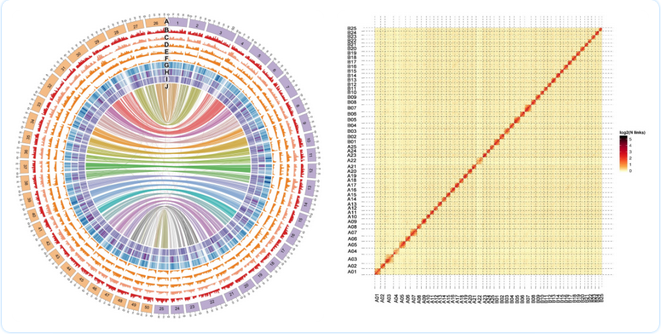
图1 金鱼A亚基因组和B亚基因组概况(A);金鱼Hi-C互作热图(B)
研究结果2:金鱼亚基因组揭秘
在过去的50年里,一直推测金鱼基因组是四倍体。近期研究表明,鲃亚科Barbinae (2n = 50) 中的鲤鱼科可能是金鱼最接近的二倍体祖先,进而形成这样一个假说:鲃亚科(Barbinae)可能是二倍体金鱼谱系的祖先种,研究中进一步进行了6个代表性的二倍体鲃亚科物种(包括条纹小鲃,大鳞高须鱼,长臀鲃,黑鳍袋唇鱼,施氏魮和裂峡鲃)的全基因组鸟枪测序并进行基因组组装。金鱼染色体可清晰分为两个亚基因组,通过reads多序列比对,发现金鱼来自两个祖先世系,其中一个是鲃亚科共有的,进一步将与鲃亚科比例高的reads所在染色体定为A亚基因组(ChrA01~A25)并将其余染色体定为B亚基因组 (ChrB01~B25)(图2)。此外,基于1个核基因的系统进化分析,即结缔组织生长因子like基因在二倍体金鱼中仅有1个拷贝,但在四倍体金鱼中含有2个拷贝,结果显示鲃亚科物种的同源基因与金鱼A亚基因组同源基因聚类到一起(斑马鱼和鲤鱼作外群)。研究结果表明,A亚基因组可能起源于鲃亚科内的祖先物种,而B亚基因组的二倍体祖先可能已经灭绝或起源于未知的鲤科鱼类谱系,推测金鱼的起源可能是由于异源多倍化事件。金鱼的A和B亚基因组分别包含28,133和26,141个基因。同时通过金鱼10个不同组织的RNA-seq测序,探索了A和B两个亚基因组之间的差异表达,发现同源基因在两个亚基因组之间表现出不对称的表达模式,而全基因组范围内的表达水平总体上更偏好A亚基因组。
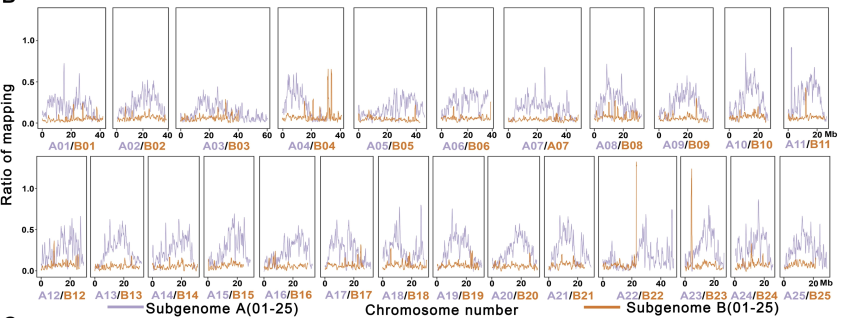
图2 鲃亚科条纹小鲃Puntius semifasciolatus Reads与金鱼的比对效率
研究结果3:金鱼基因组进化研究
在同源染色体对ChrA01/ChrB01, ChrA04/ChrB04, ChrA05/ChrB05, ChrA09/ChrB09, ChrA11/ChrB11和ChrA15/ChrB15上出现了大片段倒位,尽管整体共线性很强。这些大的片段倒位可能是异源四倍体形成的原因之一。进一步发现,金鱼两个亚基因组(Ks= ~0.17)与鲤鱼(Ks= ~0.21)之间的分歧时间(13.28-16.67 MYA),早于金鱼和鲤鱼之间的分歧时间(7.81〜8.77 MYA)(Ks =〜0.10)(图3 A)。此外,文中假设全基因组复制(WGD)事件发生在金鱼和鲤鱼的共有谱系中,因此它们可能具有相同数量的染色体(2n = 100),染色体基数(2n=50)是二倍体鲤亚科成员的两倍,暗示这两个种都是四倍体。同时,发现斑马鱼于~ 32.8-36.8 MYA,与鲫鱼、鲤鱼和草鱼由共同的祖先(LCA)分化而来(图3 B)。
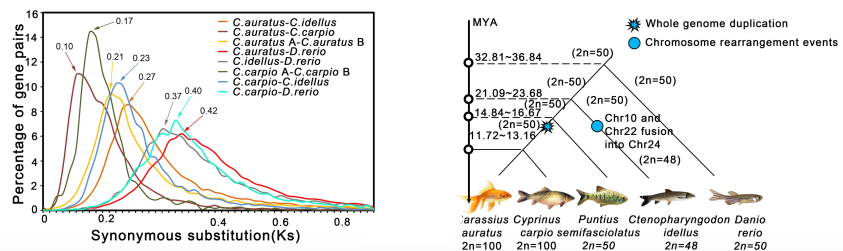
图3 金鱼、鲤鱼、草鱼、条纹小鲃和斑马鱼的全基因组复制事件(A)和系统进化分析(B)
研究结果4:金鱼群体进化研究
通过201个个体(16只野生鲫鱼和185只代表性金鱼变种,平均测序深度12.5 x)的系统发育树构建发现普通金鱼与鲫鱼亲缘关系最近(图4 A-C),进一步结合系统发育树和PCA分析确定各金鱼变种的系统分化关系,区分普通金鱼和卵金鱼(无背鳍)和温金鱼(有背鳍),无背鳍金鱼和有背鳍金鱼进一步分为不同的亚群。
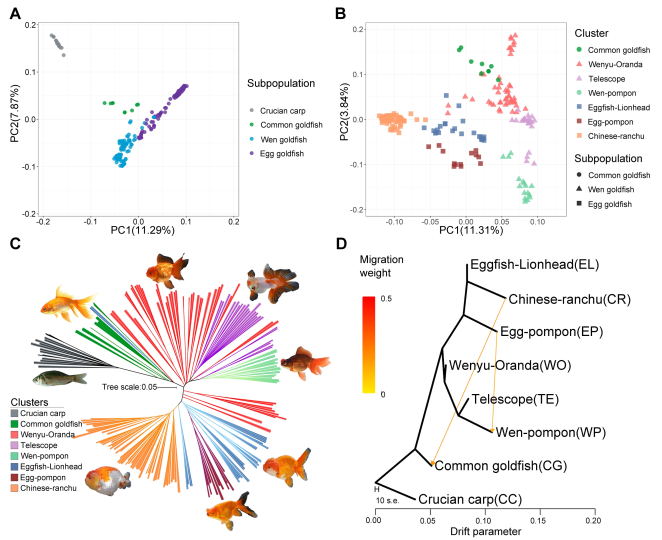
图4 金鱼群体结构分析
LD及群体选择分析支持了前期的假设,即金鱼驯化过程中种群遗传瓶颈效应的存在和强烈的人工选择(图5 A)。进一步的遗传多样性分析发现,由于人工选择的结果,从鲫鱼和普通金鱼的驯化过程中遗传多样性的增加,表明了从鲫鱼驯化后,金鱼重要遗传变异的积累(图5 B);33个金鱼个体和16个野生鲫鱼个体的群体选择分析发现了50个受选择区域(25.2 Mb区间内,共计946个基因)(图5 C),940个候选基因中大部分与形态发生,色素沉着,行为,免疫反应/传染病,能量代谢和对激素的反应有关;进一步通过以上201个个体的选择性清除分析发现393个基因受选择,其中21个代表性候选基因中,有13个与在斑马鱼中存在敲除系的基因同源,这些敲除系显示与行为相关的突变表型(pcdh15a和agrp),眼睛大小减小(ndufs5),细胞迁移(nav3和zap70)或脑大小减小(aurka)。
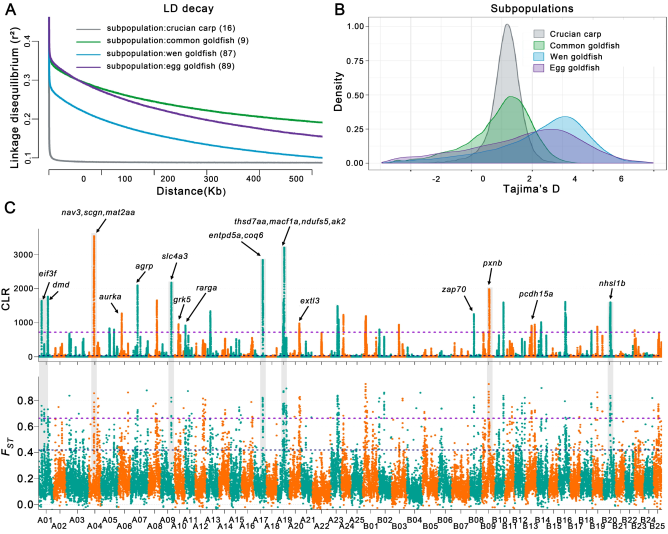
图5 金鱼群体遗传多样性和群体选择分析
研究结果5:金鱼表型性状GWAS关联分析
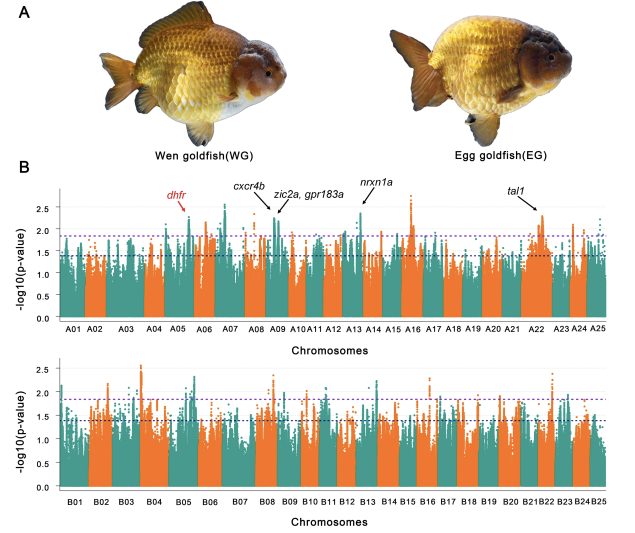
背鳍作为金鱼一重要的表型特征,在不同变种中形态不一,背鳍是区分Wen金鱼和Egg金鱼的显着特征,背鳍的缩小是Egg金鱼进化的关键特征。通过Wen金鱼和Egg金鱼杂交后代F1背鳍性状分离比,暗示了了背鳍特征受多个基因位点控制,可能具有母体遗传效应。对96只有背鳍和87只无背鳍金鱼的全基因组关联分析GWAS研究分析发现跨越13条染色体上的8.96 Mb区域上的378个基因与背鳍性状相关(图6),85.2% (322)个基因位于A亚基因组,大部分位于五条染色体上,包括ChrA09,ChrA07,ChrA22,ChrA16和ChrA05,比位于B亚基因组上的背鳍相关基因的数量高得多,这表明背鳍相关基因不对称的亚基因组分布,GO和KEGG分析表明,该性状的候选基因可能参与了“细胞表面受体信号传导”途径,“信号转导”,“跨膜转运”,“骨骼系统发育”和“初级代谢过程和器官氮化合物代谢过程”。进一步通过与斑马鱼直系同源基因基因敲除品系表现出的突变表型分析发现,二氢叶酸还原酶(dhfr)可能在某种程度上与鱼鳍是否存在相关,该基因可能在金鱼人工选择过程中,在有无背鳍这一表型中发挥重要作用。
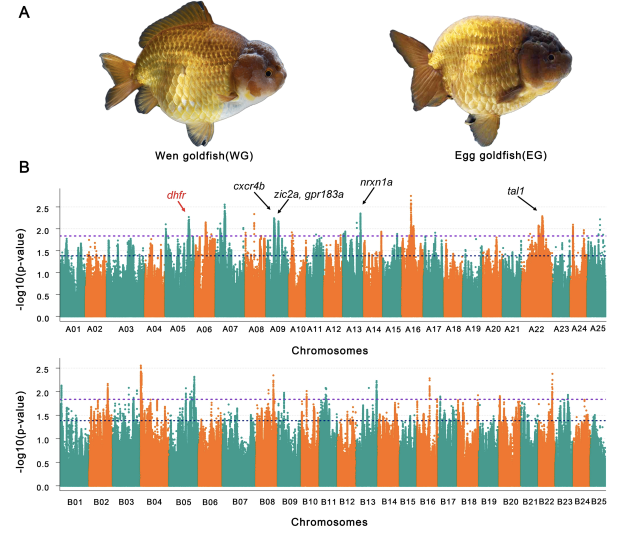
图6 金鱼背鳍相关性状GWAS关联分析





 京公网安备 11011302003368号
京公网安备 11011302003368号