

2022年6月,华中农业大学园艺植物生物学教育部重点实验室邓秀新院士团队在National Science Review上发表了题为“Structural variation and parallel evolution of apomixis in citrus during domestication and diversification”的研究论文,首次揭示了柑橘中不同属间无融合生殖性状存在平行进化,回答了现代栽培柑橘广泛存在无融合生殖的遗传机制,对柑橘育种研究具有重要启示。百迈客有幸参与本项研究,完成其中PacBio、Hi-C测序等工作。
研究背景
我国是柑橘的起源地之一,栽培品种类型繁多,柑、橘、橙、柚、葡萄柚、柠檬、金柑、杂柑等都有规模化的栽培。柑橘广泛存在无融合生殖的特性,导致后代基因型和母本保持一致,阻碍了柑橘的杂交育种进程。因此,研究柑橘无融合生殖对于加速柑橘遗传改良具有重要意义。此外,无融合生殖还被运用于作物的杂种优势固定,具有重要应用前景,对我国种业发展具有重要意义。柑橘是研究孢子体无融合生殖的模式材料,解析柑橘无融合生殖的遗传机制,能够为作物杂种优势固定提供新的思路。
柑橘中的金柑属在我国具有悠久的栽培历史,山金柑 (Hongkong kumquat, Fortunella hindsii) 是金柑属的一种常绿野生果树,具有无融合生殖和有性生殖两种类型,是研究无融合生殖的良好材料。
主要研究结果
柑橘基因组组装与结构变异
为了解析柑橘珠心胚的进化、遗传和分子机制,作者对山金柑自交系Sy3-45进行了基因组组装。最终完成了323.72 Mb(contigN50=9.77 Mb)的高质量基因组(图1a),其连续性比之前的版本提高了4.4倍。BUSCO评估(97%)、共线性评估,均显示本次构建了高质量的柑橘基因组。通过对不同起源、不同生殖模式柑橘的半合子基因数量进行评估,发现半合子基因数量与生殖模式有关,克隆和无融合生殖作物的半合子基因数量最高,而自交作物的半合子基因数量较低。
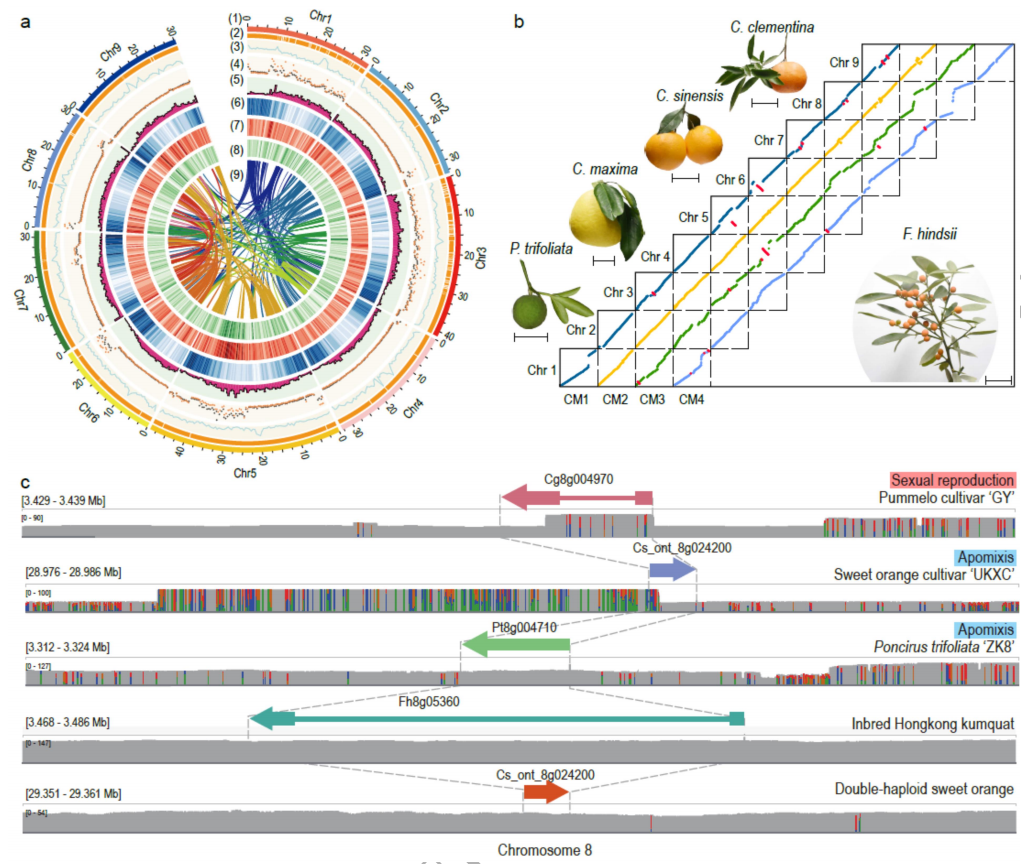
图1 山金柑基因组特征与基因组的比较
柑橘无融合生殖表型的多起源
收集了234份柑橘亚族的材料全基因组测序数据(其中46份是该研究新测的山金柑种质),这些数据中包括介于杂交起源的混合类群(即葡萄柚、甜橙、柠檬和酸橙)。系统发育分析表明,生殖模式在多种情况下从完全有性生殖向无融合生殖和再回到有性生殖(图2)。无融合生殖表型在柑橘中广泛分布,研究表明无融合生殖可能在该类群中有多个起源。对群体结构分析区分的12个类群进行全基因组的基因深入分析,发现基因渗入在全基因组广泛存在。但聚焦到无融合生殖区段后发现,渐渗并不能解释柑橘属和金柑属无融合生殖的发生。
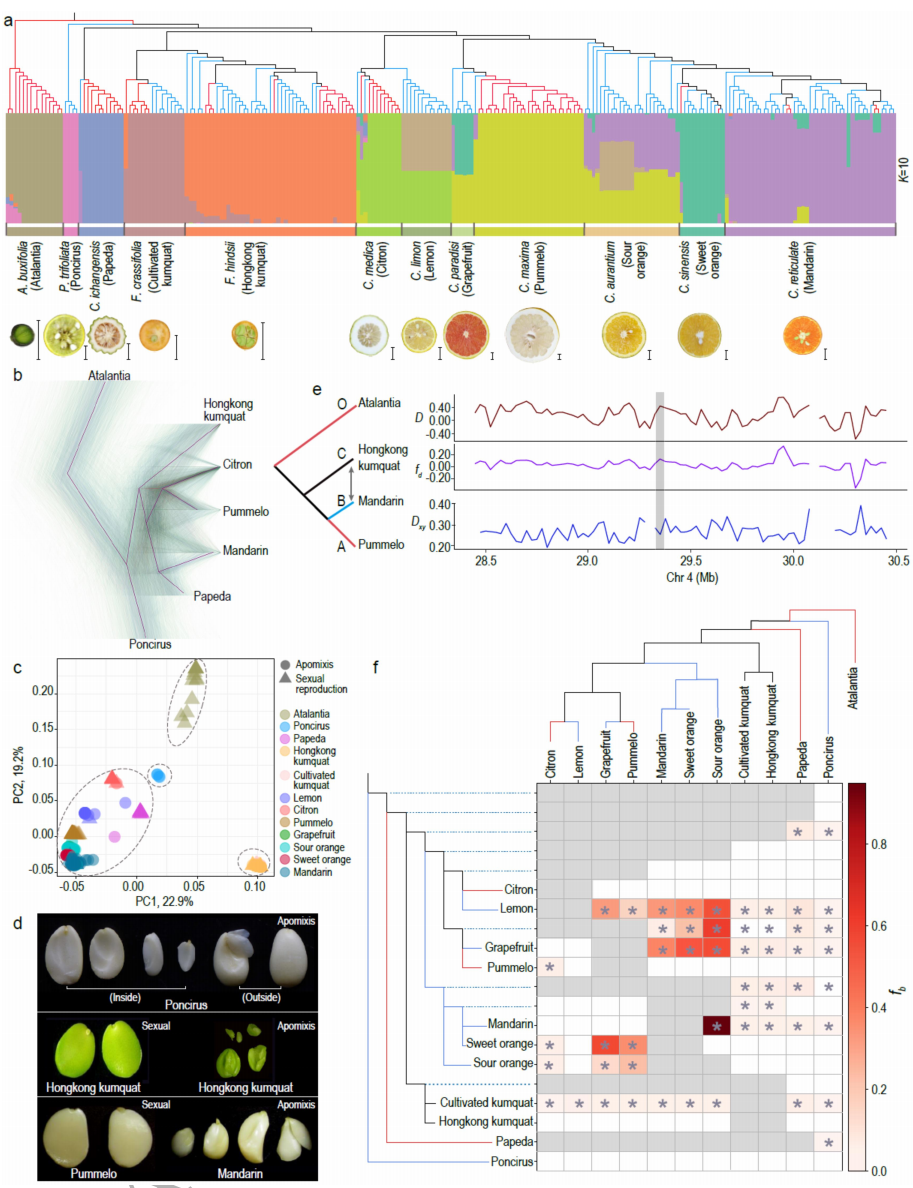
图2 柑橘群体进化研究
无融合生殖和有性生殖群体的结构和有害负担
柑橘通过珠心进行无融合生殖,保留了母体基因型,这可能导致基因组中有害突变和结构变异(SVs)的积累。从234份材料中共鉴定出139,241个可靠的SVs,其中64.2%为缺失,16.6%为重复,15.5%为易位。山金柑的SV数量最高,可能与有效种群大小减少有关(图3a)。具有无融合生殖的杂交群体的杂合有害变异约为有性生殖群体的1.45倍。随后提出了杂种无融合生殖将SVs隐藏在杂合子状态的假说,这可能主要是隐性选择造成的危害。因此,有性生殖的杂种可能会迅速以隐性纯合的方式暴露有害变异。这个假说解释了为什么无融合生殖和有性生殖的天然杂交样本中观察到的自然杂种通常是无融合生殖的。有害突变的模式与SVs的模式一致,即与有性生殖群体相比,无融合生殖群体的隐性有害负担较低(7.39%),杂合子SVs较多(约2.62倍)。在隐性模型下,从无融合生殖到有性生殖的生殖模式转变导致种群适应度在转变后立即下降13.48±0.87%,但随后恢复(图3i)。相比之下,保持无融合生殖的种群保持种群适合度。
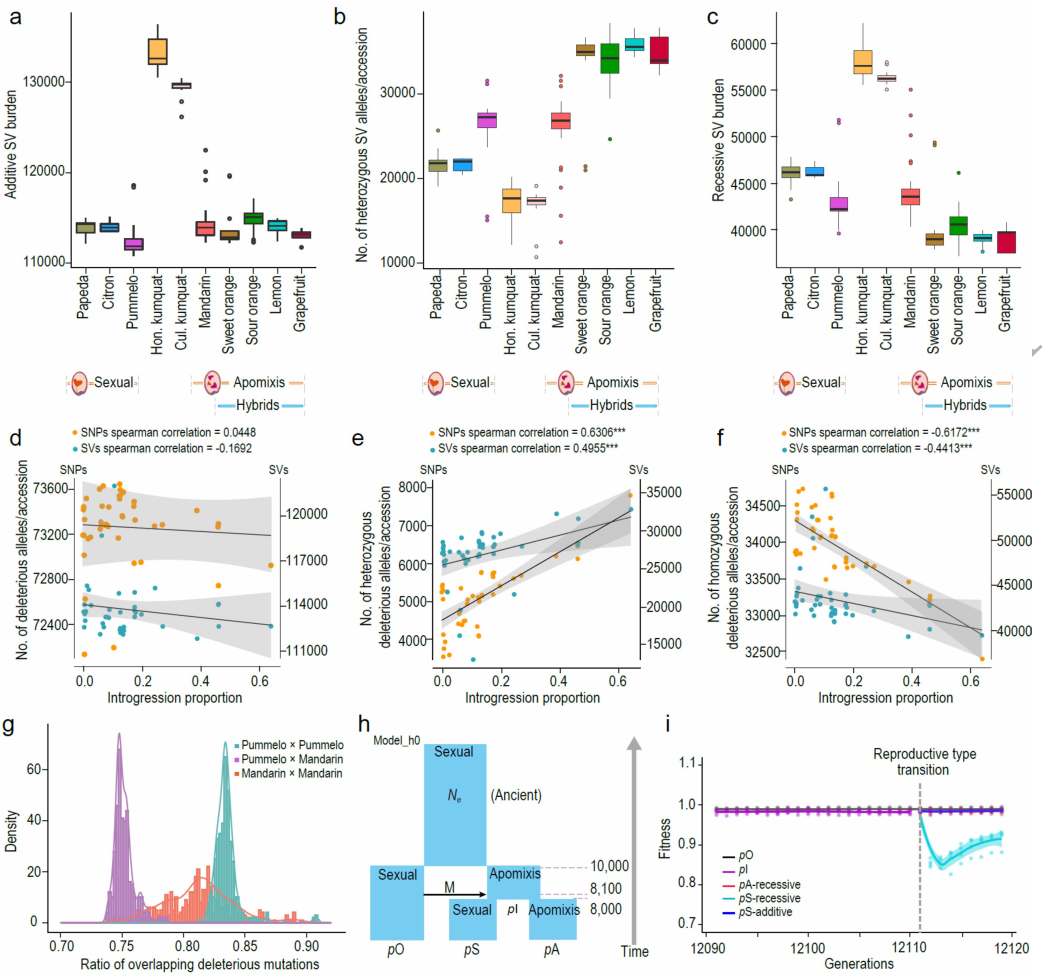
图3无融合生殖和有性生殖群体中的结构变异(SVs)和有害突变
同一位点有助于金柑属和柑橘属的无融合生殖
山金柑具有无融合生殖和有性生殖两种类型,为了定位山金柑无融合生殖的潜在位点,作者利用无融合生殖的山金柑系“DB”与两个有性生殖的山金柑“PN”和“PN03”作为母本杂交构建了一个分离群体(图4a)。通过ΔSNP指数及KASP标记等方式进一步将相关位点缩小到chr4上313 Kb区域。基于313 Kb区域的系统发育学及全基因组fb统计表明,至少有一些山金柑的无融合生殖是通过基因渗入产生的。进行序列比对以进一步确定与山金柑(313 Kb)、柑橘(380 Kb)和柚子(80 Kb)基因组共享的无融合生殖基因组区域中珠心胚发生和发育相关的主要原因(图5a)。这三个区域之间的最小重叠包含FhRWP基因,这是一种RWP-RK域转录因子,与据报道与有助于柑橘无融合生殖的CitRWP基因同源。
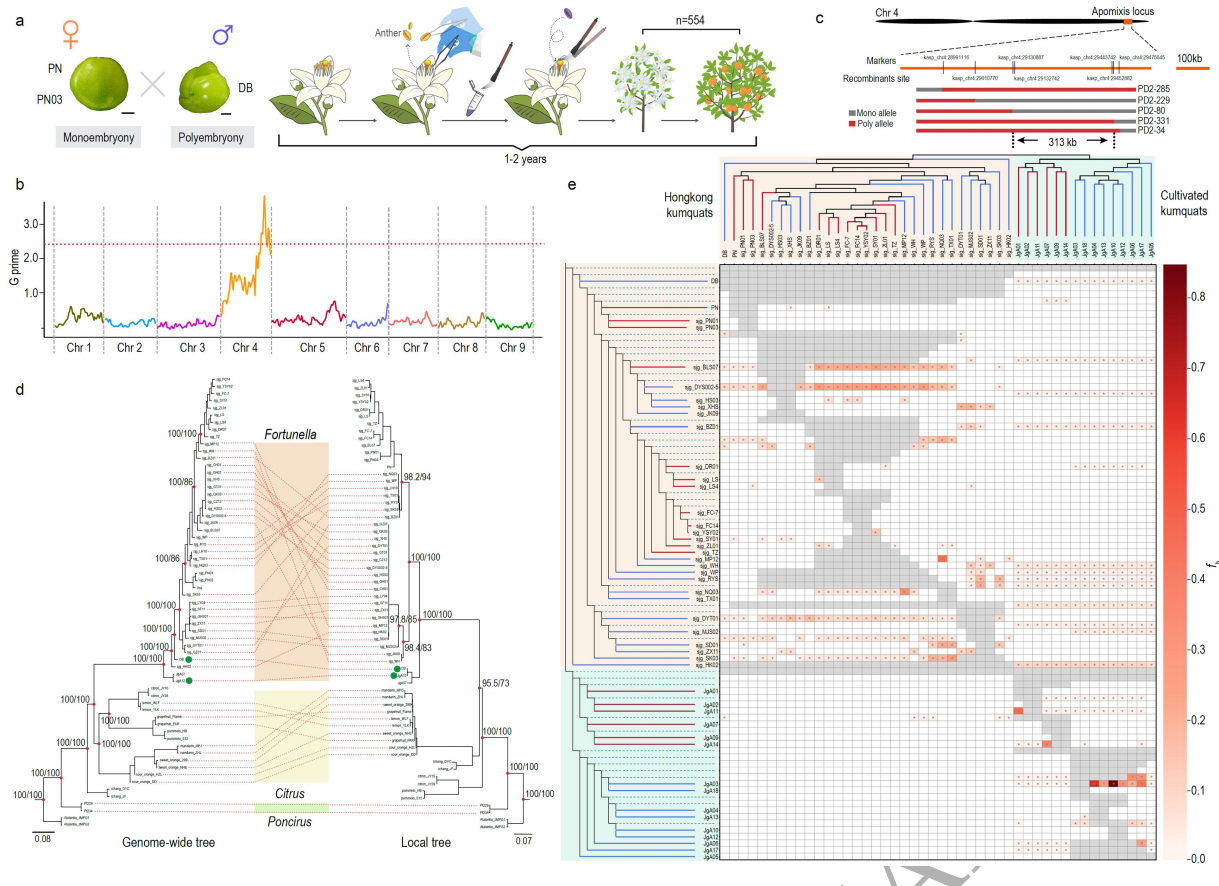
图4 有性生殖向无融合生殖过渡的候选位点
柑橘亚族无融合生殖的杂合MITE插入
为了研究柑橘无融合生殖决定区的遗传变异模式,基于单倍型构建了313 Kb区域的系统发育树(图5b)。结果表明,柑橘存在多种无融合生殖现象。同一个体的两个单倍型并不总是聚集在同一分支中,这表明不同群体中无融合生殖的起源不同。前期研究表明,在CitRWP基因启动子中插入的MITE与无融合生殖相关。作者将PCR和循环一致性测序(CCS)相结合,以阐明FhRWP启动子的复杂结构。结果证实,MITE的插入与无融合生殖和无融合生殖单倍型密切相关。进一步研究表明,不同数量的MITE及其不同的序列表明无融合生殖等位基因可能起源不同;MITE插入在杂合子状态下非常常见,这可能与无融合生殖品种的单倍型差异有关。
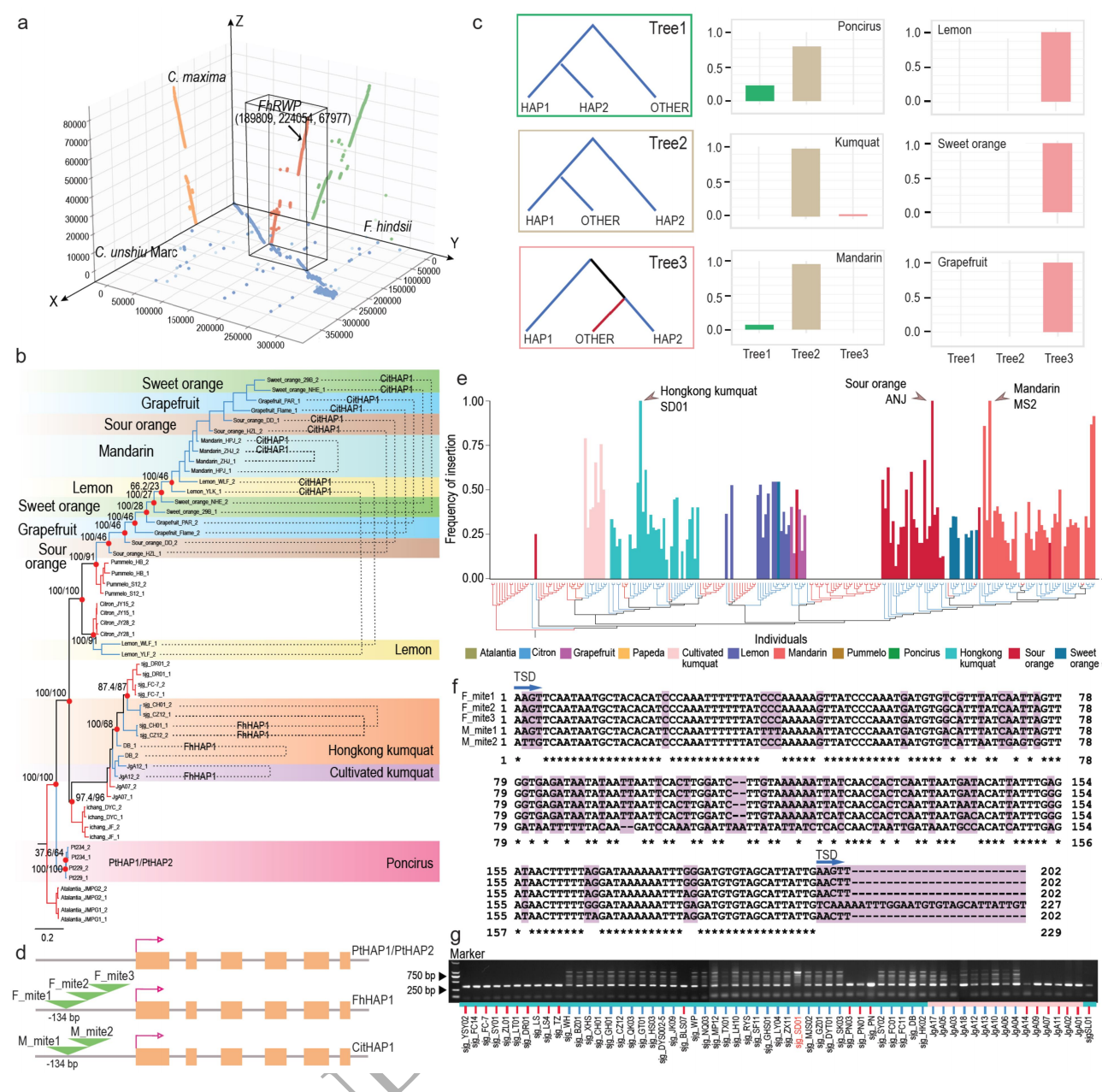
图5 山金柑和柑橘中的杂合MITE插入
与FhRWP中MITE相互作用的因素
通过细胞学分析珠心胚的起始过程,无融合生殖山金柑的胚珠在开花时明显,但在有性生殖山金柑中不明显(图6a)。转录组分析表明,在开花前7天和开花后7天两个时期,有MITE插入的无融合生殖单倍型中均检测到FhRWP表达,但在无MITE插入的有性生殖材料中未检测到表达(图6b)。在开花后0、3、5、10和20天的五个发育阶段的RT-qPCR检测证实了这一结果。
甲基化及酵母单杂分析进一步的探索了FhRWP基因表达调控的相关因素,鉴定了一种转录因子FhARID(Fh3g37720),位于与无融合生殖区不同的染色体上,该转录因子编码一个富含AT的相互作用域的蛋白质,该蛋白质结合到FhpMITE右侧片段(图6h)。FhARID在无融合生殖的山金柑胚珠中特异表达。为了研究MITE类序列的数量是否对FhARID诱导的FhRWP表达激活很重要,作者构建了具有一个、两个和三个pMITE序列拷贝的启动子,并通过瞬时转化烟草进行了双荧光素酶测定。结果表明,FhARID结合pMITE序列并激活连锁报告基因表达,但pMITE序列的数量在FhARID介导的激活中没有显示出显著差异。
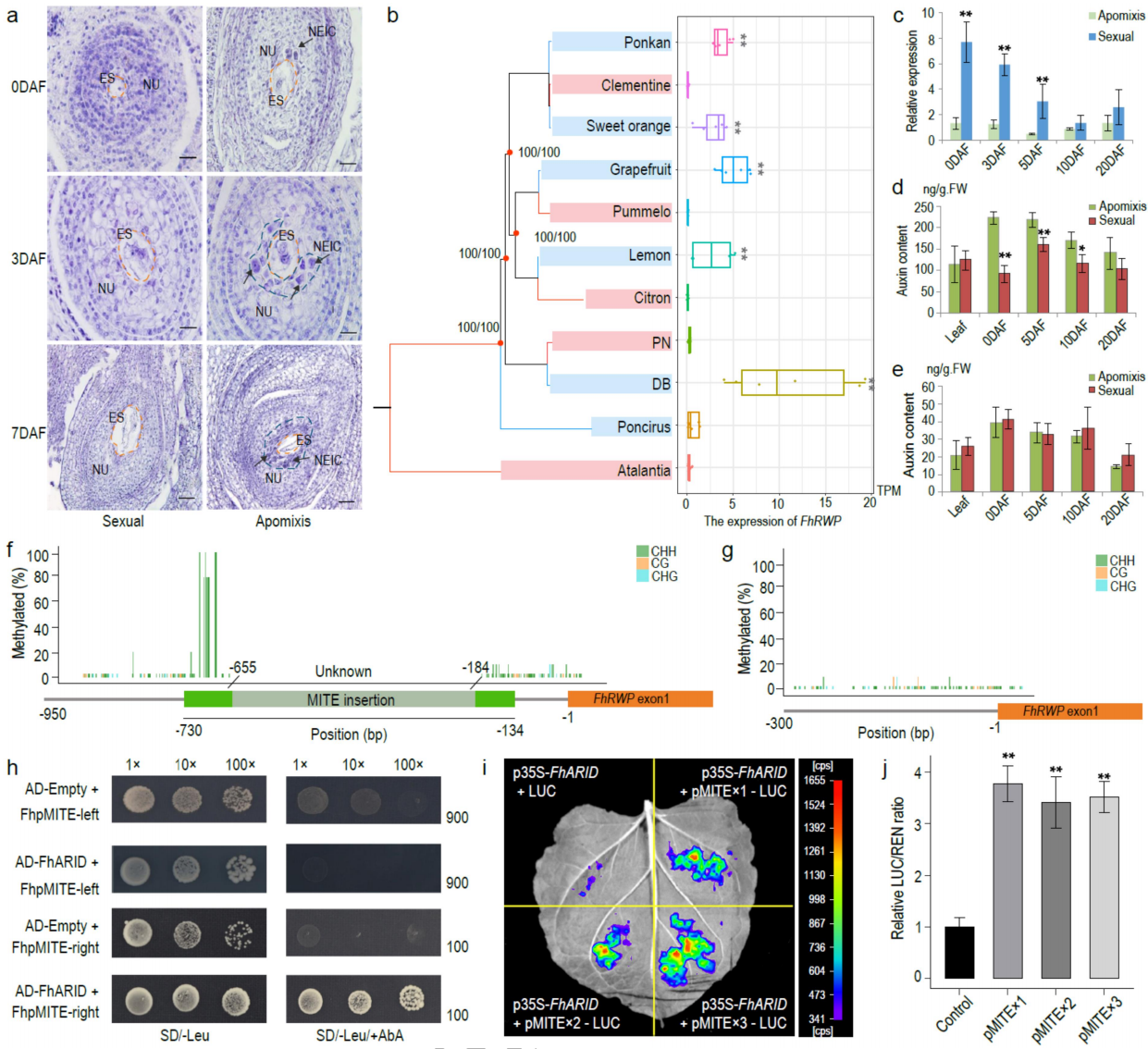
图6 无融合生殖调控的分子特征
研究小结
该研究团队利用山金柑品系Sy3-45为材料进行了高质量染色体级别基因组的组装和遗传群体构建,实现了金柑属无融合生殖关键基因的定位。结合柑橘不同属材料的全基因组重测序数据,进一步解析了金柑属和柑橘属无融合生殖表型的平行进化机制。该研究发现,杂交起源的柑橘群体能够通过无融合生殖来保持杂交优势,并导致无融合生殖的关键MITE插入以杂合子形式存在,首次提出了无融合生殖在柑橘分化和驯化过程中的重要作用。
如果您的项目需要PacBio、Hi-C测序服务,欢迎点击下方按钮联系我们,我们将免费为您进行文章思路方案设计。





 京公网安备 11011302003368号
京公网安备 11011302003368号