

 文章标题:Molecular and spatial signatures of human and rat corpus cavernosum physiopathological processes at single-cell resolution
文章标题:Molecular and spatial signatures of human and rat corpus cavernosum physiopathological processes at single-cell resolution研究背景
阴茎勃起是一个复杂的生物学过程,整个过程需要神经、内分泌、血管和阴茎海绵体组织精密调节、协调完成。海绵体损伤可引起多种疾病,包括影响了5%~22%男性的阴茎勃起功能障碍(ED)。
ED不仅影响患者身心健康以及家庭和谐,还可能是心血管疾病的先兆。虽然目前已经知道在勃起发生和维持中不同功能区域的作用,但以往研究大多都依赖于解剖观察、影像学检查,以及少数基因或蛋白质的表达模式分析,缺乏全面的分子水平分析。
此外,多数ED相关研究是基于大鼠模型的,但当前对于大鼠和人类间阴茎海绵体(CC)的解剖学和信号网络差异认识尚不清晰。因此,为了更好的使用大鼠模型开展研究,有必要深入研究大鼠阴茎海绵体的结构/功能调控与人类的异同。
材料方法
研究材料:4例阴茎癌患者,3例健康志愿者,10例DMED(糖尿病性勃起功能障碍)患者;4只健康大鼠,6只DMED大鼠模型。
研究方法:Masson/H&E染色(n=7),scRNA-seq(n=12,3例健康人类CC/3例DMED人类CC,3例健康大鼠CC/3例DMED大鼠CC),空间转录组技术(BMKS1000,n=3,1例阴茎肿瘤患者、1例勃起功能正常雄性大鼠以及1例DMED模型大鼠)等。
验证实验:包括油红O/尼罗红/菲律宾链霉菌染色、不同硬度ECM条件下培养的FB细胞形态信息及bulk RNA-seq数据、免疫荧光、超声弹性成像。
研究结果
1.人类和大鼠CC细胞和空间特征
通过scRNA-seq数据,研究者在人类CC(阴茎海绵体)中鉴定出7种类型细胞,仅在大鼠CC中发现一小部分中性粒细胞和B细胞。使用scRNA-seq数据对空间转录组数据进行注释(CellTrek),研究者得到了人类和大鼠CC样本空间细胞分布图谱(level3,20μm分辨度),发现虽然人类和大鼠CC中的细胞类型相似,但每种类型细胞比例、空间分布特征存在异质性。
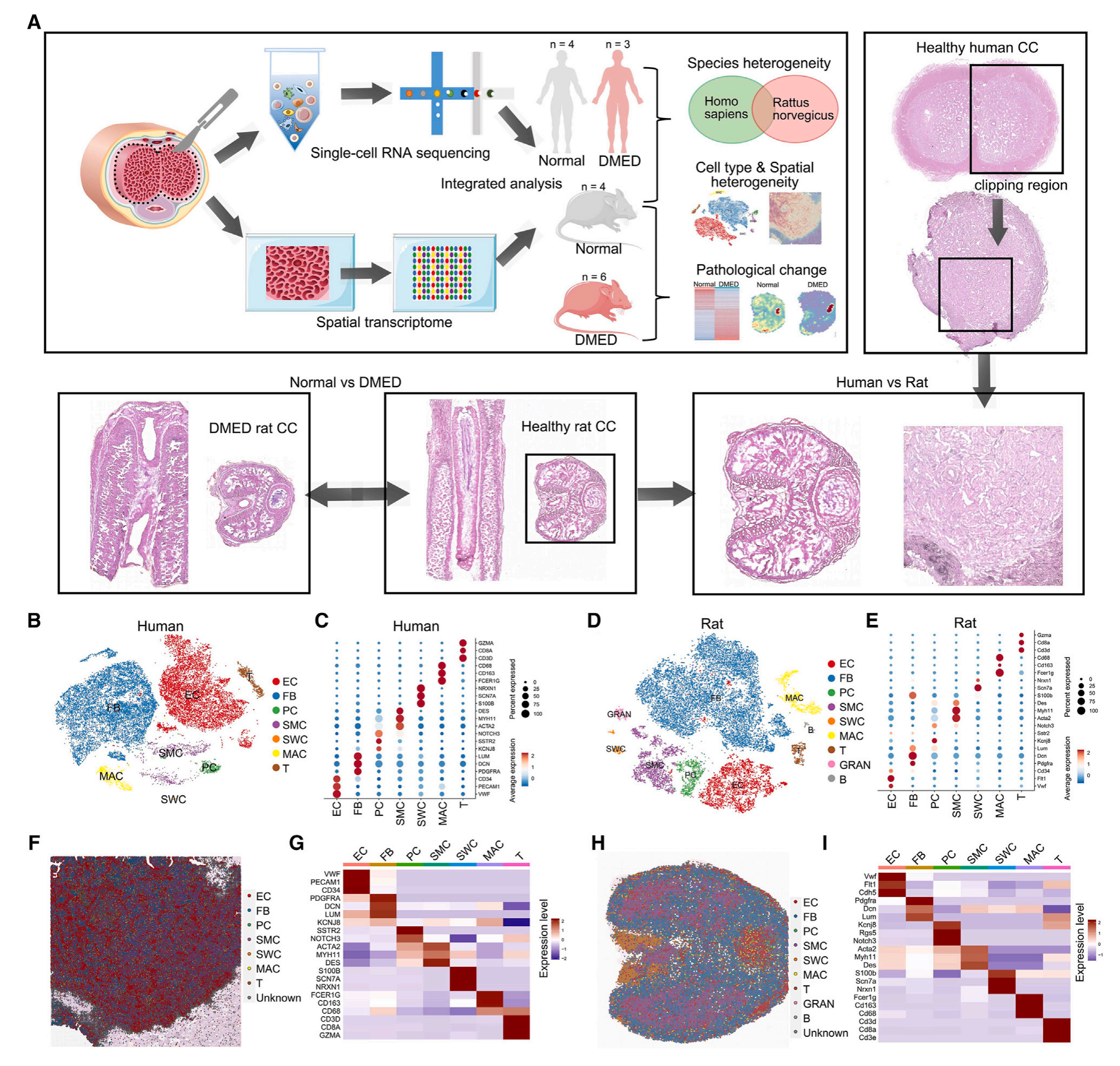
图1-通过scRNA-seq和空间转录组技术得到的人类,以及正常/DMED条件下大鼠的CC细胞全局表达谱
2.人类和大鼠CC组织中的空间异质性
作为一种特殊的血管窦结构,当前已初步认识到CC上不同区域的类型不同。得益于空间转录组技术,研究者发现人类CC组织中SVG(空间差异基因)top(如CCL18、PLA2G2A、C3)主要在不同类型细胞中表达,但有一些表现出空间分布特异性的基因并没有在scRNA-seq数据中表现出细胞类型特异性;人类CC组织3个代表性区域中,不同区域的细胞比例不同,如区域I(海绵体动脉区域)EC(内皮细胞)占比更高,而区域III(白膜附近区域)包含更少的FB(成纤维细胞)但SMC(平滑肌细胞)丰富。大鼠CC组织也表现出相似的情况(大鼠CC中缺乏梳状中隔和明显的海绵体动脉,研究者根据人类CC代表性区域的空间位置,在大鼠CC中也划分出可比较的3个代表性区域)。
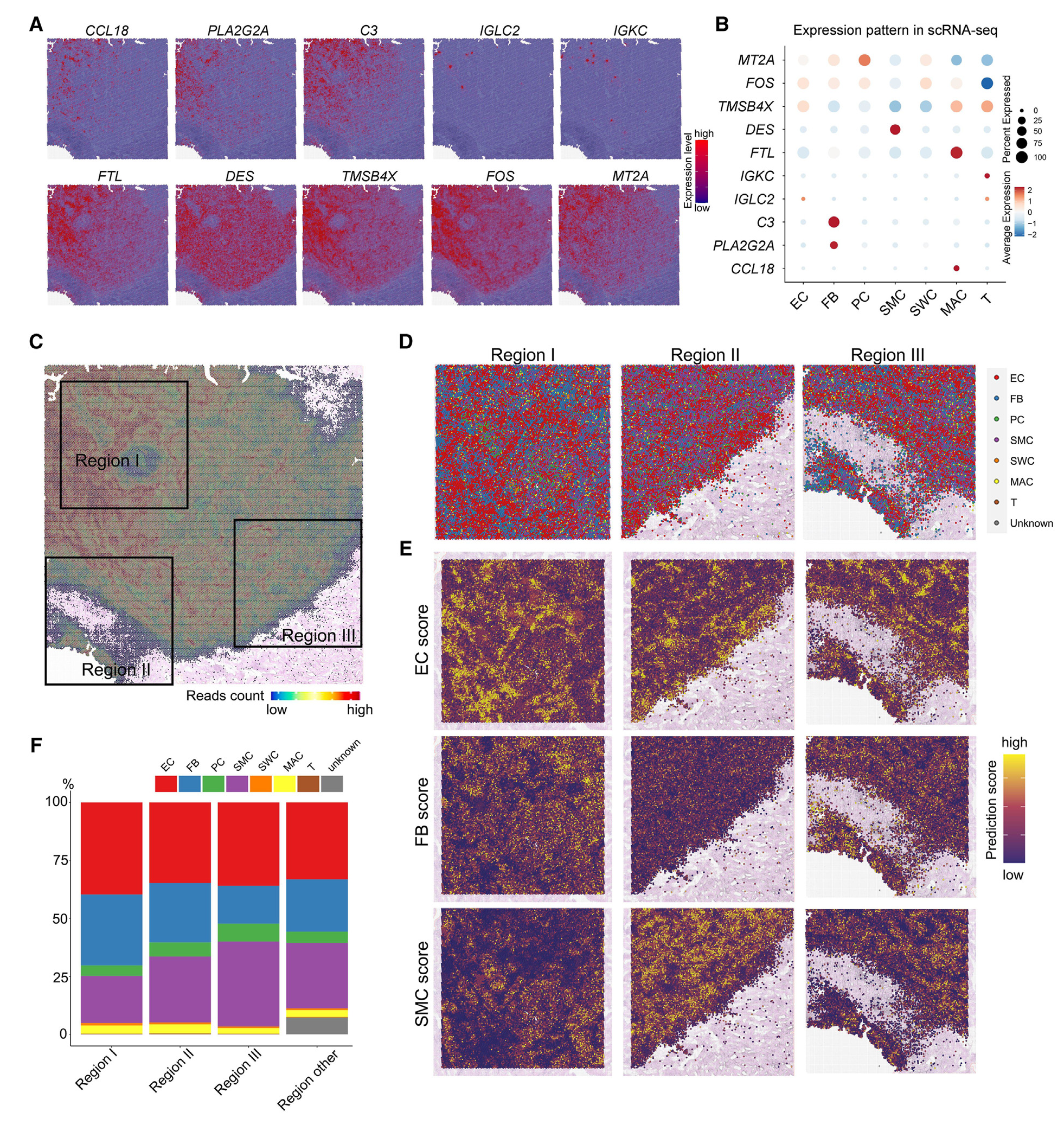
图2-人类CC组织的转录特征和空间异质性
3.根据scRNA-seq数据比较人类和大鼠CC中的细胞类型
多数勃起调控研究是基于大鼠模型的。为了理解大鼠和人类间CC微环境的异同,研究者通过分析scRNA-seq数据中的人—大鼠同源基因,发现虽然人类和大鼠物种不同,但CC中相同类型细胞仍可以聚成同一簇,进一步分析发现人类和大鼠CC中相同类型细胞的转录相似性>70%且正相关,表明在多数情况下,使用大鼠模型模拟、研究人类CC微环境的转录调控是可接受的。然而,人类和大鼠CC中相同类型细胞的DEGs(差异表达基因)top并不一致,提示当研究者更关注一个或几个基因或通路时,使用大鼠模型要特别注意。

附录图7-scRNA-seq数据评估得到的大鼠和人类CC间相似性和差异性
4.比较人类和大鼠CC空间转录组全局
通过分析空间转录组数据中的人—鼠同源基因,研究者找到人类和大鼠共有的459个top SVG,大多数SVG并不是细胞类型特异的,其中EC、FB和SMC是表达这些SVG最多的3种类型细胞。富集分析结果显示,基础生物学过程如“translational initiation”、“extracellular structure organization”以及“protein targeting”,在人类和大鼠CC组织的空间转录组数据中是保守的;人类特异SVG主要与炎症响应有关,而这些SVG的大鼠同源基因主要与代谢过程有关。

图3-比较大鼠和人类CC微环境中的空间差异基因和信号通路
5.人类和大鼠CC微环境中的细胞—细胞互作分析
为了研究、比较人类和大鼠间CC微环境的复杂信号网络,研究者使用scRNA-seq数据进行CellChat分析,发现大鼠CC中的细胞互作数量虽然小于人类,但细胞互作强度是相似的。基于这些互作的表达模式,研究者进行简单的分类,发现虽然胞外基质相关的互作整体强度与细胞类型间配体-受体强度相当,但两者基因组成是有差异的;血管生成相关的互作整体强度在人类和大鼠中没有差异,但基因组成存在差异,如大鼠CC微环境中是主要是VEGFB表达,而人类CC微环境中高表达的是VEGFA和IGF1。
空间转录组数据显示,VEGF配体主要集中富集在人类CC中的海绵体动脉(区域I)附近,而在蛋白质水平,VEGFA和IGF1并不共定位。大鼠中VEGF和IGF基因的空间分布模式与人类的相似,但蛋白质水平上区域I中存在高浓度的VEGFA和IGF1。此外,免疫相关的互作在人类CC与大鼠CC中表现出相似或者更高的整体信息流,但一些配体亚型在大鼠中高表达。
这些scRNA-seq数据的分析结果与基于空间转录组数据SVG的富集分析结果高度一致。
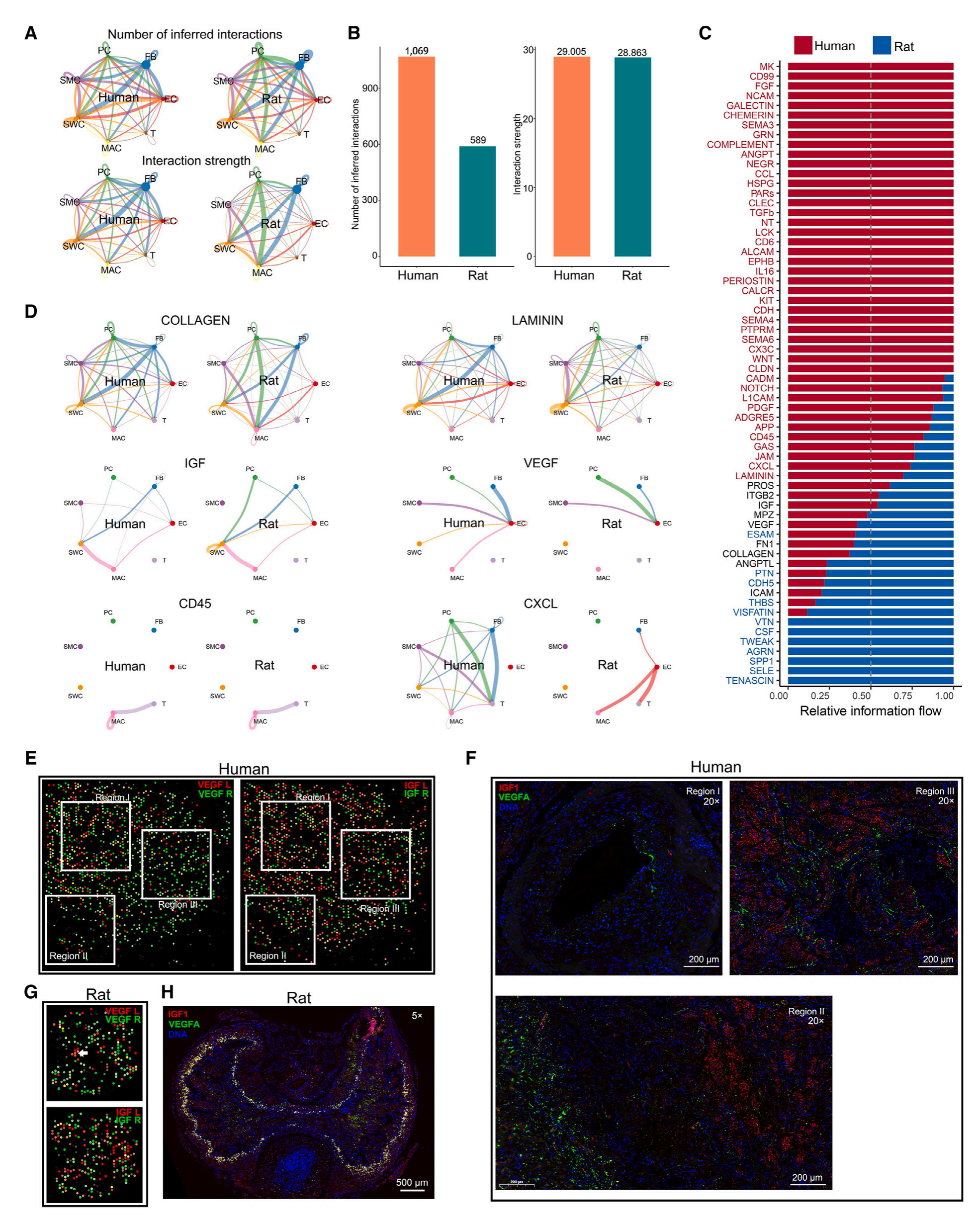
图4-配体-受体对分析显示人类和大鼠CC组织中的细胞—细胞通讯
6.EC表现出显著的空间异质性和物种差异
根据以往的解剖观察以及scRNA-seq数据分析,可知CC中包含3种类型EC(内皮细胞),包括GJA5+ EC(动脉内皮细胞)、SELP+ EC(静脉内皮细胞)以及KIT+ EC(海绵体窦内皮细胞)。人类CC中GJA5+ EC占比最低,主要分布在海绵体动脉区域;SELP+ EC主要分布在白膜区域;KIT+ EC广泛分布。但大鼠CC中,Selp+ EC比例更高,Gja5和Kit可能并不是有用的静脉和CC内皮细胞标志物。基于scRNA-seq数据进行GO分析,研究者发现不同类型EC富集了不同功能,多数细胞类型特异性的term(条目)在人类CC中并没有空间分布异质性,但一些特殊term在大鼠CC中表现出空间分布异质性。
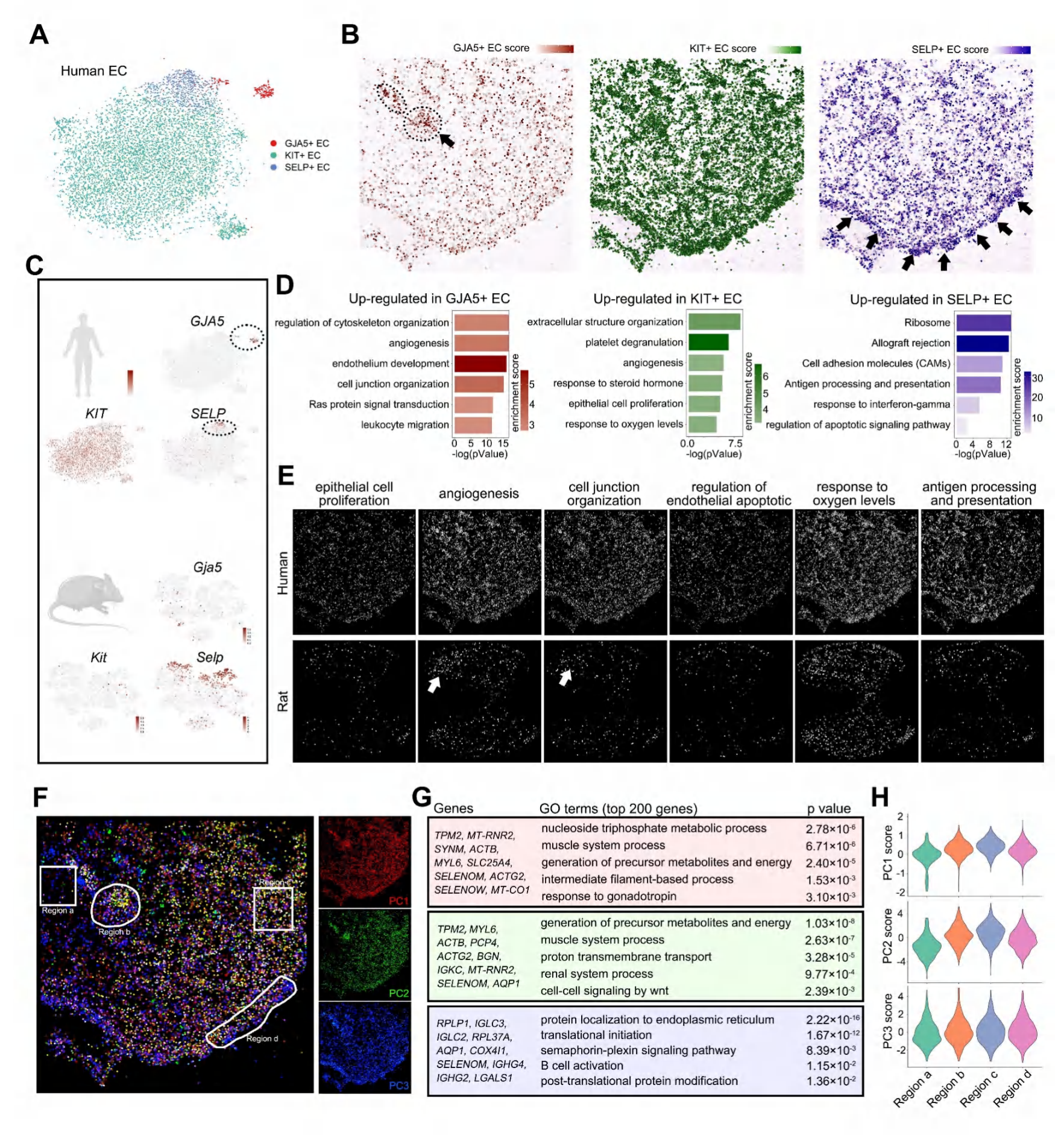
附录图10-人类和大鼠EC簇间的物种相似性和异质性
7.SMC表现出显著的空间异质性和物种差异
根据以往研究结果,研究者将CC中的SMC(平滑肌细胞)簇分为VSMC(血管平滑肌细胞)和CCSMC(海绵体小梁平滑肌细胞)。数据分析结果显示,两种类型SMC在物种间存在显著差异,其中人类CCSMC高表达DES,VSMC高表达RGS5;大鼠CCMS和VSMC可通过Igfbp2和Rgs5区分;蛋白质水平上,CCSMC与VSMC不同,后者不表达肌动蛋白或肌球蛋白;根据SMC的SVG空间分布模式,发现ACTC1/Actc1只在人类CCSMC中表达;前期研究发现ADRA2A在人VSMC表达,ADRA2C在人CCSMC中表达,空间转录组数据也验证了这一发现,但大鼠CC中Adra2a并没有表现出细胞类型特异性而Adra2c的表达未检测到,提示在这个方面大鼠并不是合适的研究模型。
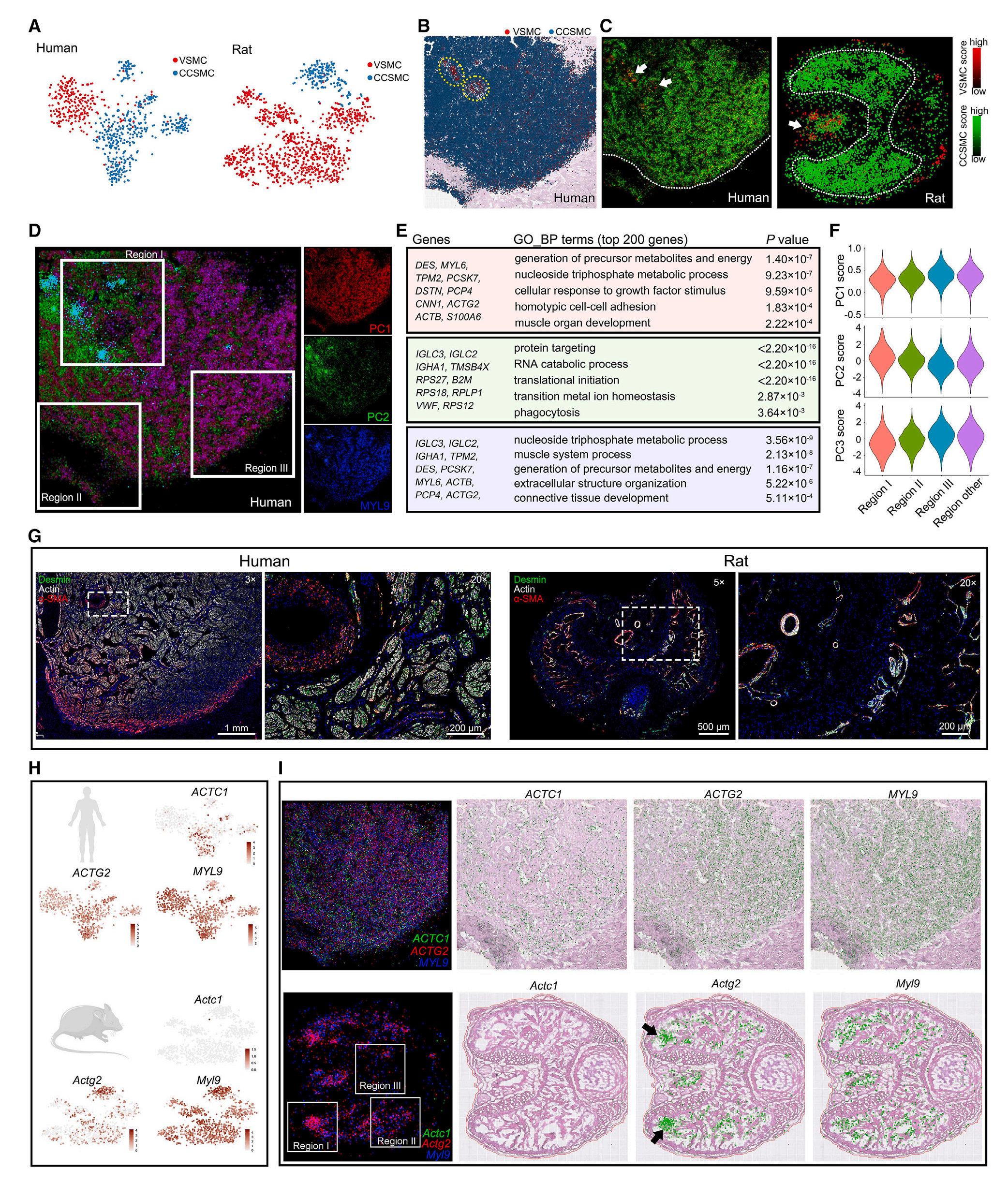
图5-人类和大鼠SMC簇间的物种相似性和异质性
8.人类和大鼠FB有相似的聚类特征和空间分布,但亚型比例不同
近期,CC的FB(成纤维细胞)成为研究热点,但对其空间和物种异质性仍不清楚。研究者根据PI16、APOE及其他载脂蛋白、COMP的表达情况,将人类CC FB分成3种亚群。空间转录组数据分析结果显示,人类CC中3种FB亚群空间分布模式不同,富集的功能也不同;大鼠CC FB表现出与人类CC FB相似的亚群和标志基因,但每种亚群的比例与人类显著不同。
APOE与脂滴转运密切相关。油红O染色显示大鼠CC中脂滴位置与Apo+ FB的空间分布高度一致,且都在白膜下富集;免疫荧光染色结果显示大多数脂滴与APOE蛋白共区域化出现且在白膜下有更高的富集,特别是在区域III(左右CC连接区域);人类CC中APOE分布特征与大鼠类似但表达更广泛。
前期研究成果显示,FB是人类CC中最强的外向信号源,在调控微环境稳态中起到重要作用。因而,FB表型转变可能与CC结构和功能变化密切相关。通路活性分析显示PI3K通路活性分值与APO+ FB有相似的空间分布模式,而TNF-α和TGF-β的空间分布模式与之相反;空间上CCSMC与FB相距较远(level13,100μm分辨度分析FB生态位),而GJA5+ EC、VSMC、SWC(Schwann细胞)和T细胞与COMP+ FB空间距离较近,表明COMP+ FB与神经和血管密切相关;CXCLs、CCN2以及C3与3种FB亚群生态位正相关,但CCN5和C7与COMP+ FB生态位负相关,GREM1不与PI16+ FB或APO+ FB相关但与COMP+ FB生态位显著正相关。
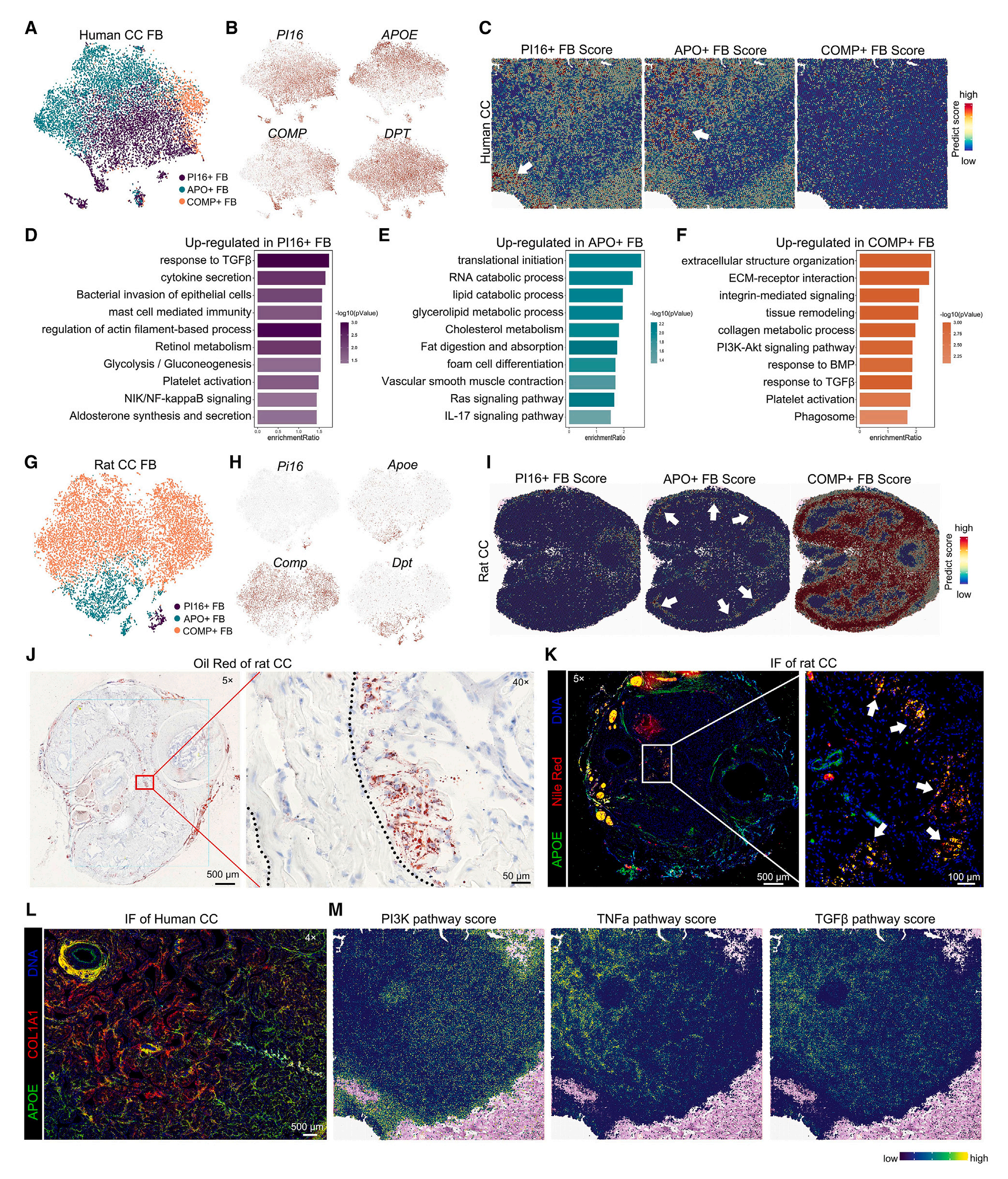
图6-CC中不同成纤维细胞亚群表现出不同的空间分布特征
9.CC中机械力信号存在空间异质性,并调控FB表型转变
以往研究表明病理条件下YAP信号诱导FB-肌成纤维细胞表型转变,但这不能完全解释生理条件下存在着的多种FB亚型,特别是有高脂质代谢活性的APO+ FB。有报道ECM(胞外基质)机械力调控多种细胞类型的脂质代谢。
本研究中,研究者发现COMP+ FB生态位显著富集了“integrin-mediated signaling pathway”和“response to mechanical stimulus”term;人和大鼠CC中ECM/细胞组成比例表现出明显的空间异质性;空间转录组数据和免疫荧光染色结果类似,靠近阴茎背神经血管束的上极区域显示出较低的纤维化水平,而远离背神经血管束的显示出较高的纤维化水平;空间转录组数据和超声弹性成像结果都表明,APO+ FB与更柔软的组织硬度有关,而COMP+ FB与更硬的组织有关;不同胞外基质硬度培养下的CC FB形态不同,bulk RNA-seq数据显示机械应激显著改变了FB的转录状态,一些参与调控脂质代谢的基因与低机械力信号有关;scRNA-seq数据分析结果显示APO+ FB分值随组织硬度增加而降低,COMP+ FB分值随组织硬度增加而增加;染色结果显示随着组织硬度增加,FB中的中性脂质和胆固醇累积降低。
综上,这些结果表明机械力信号直接参与调控FB的表型转变。
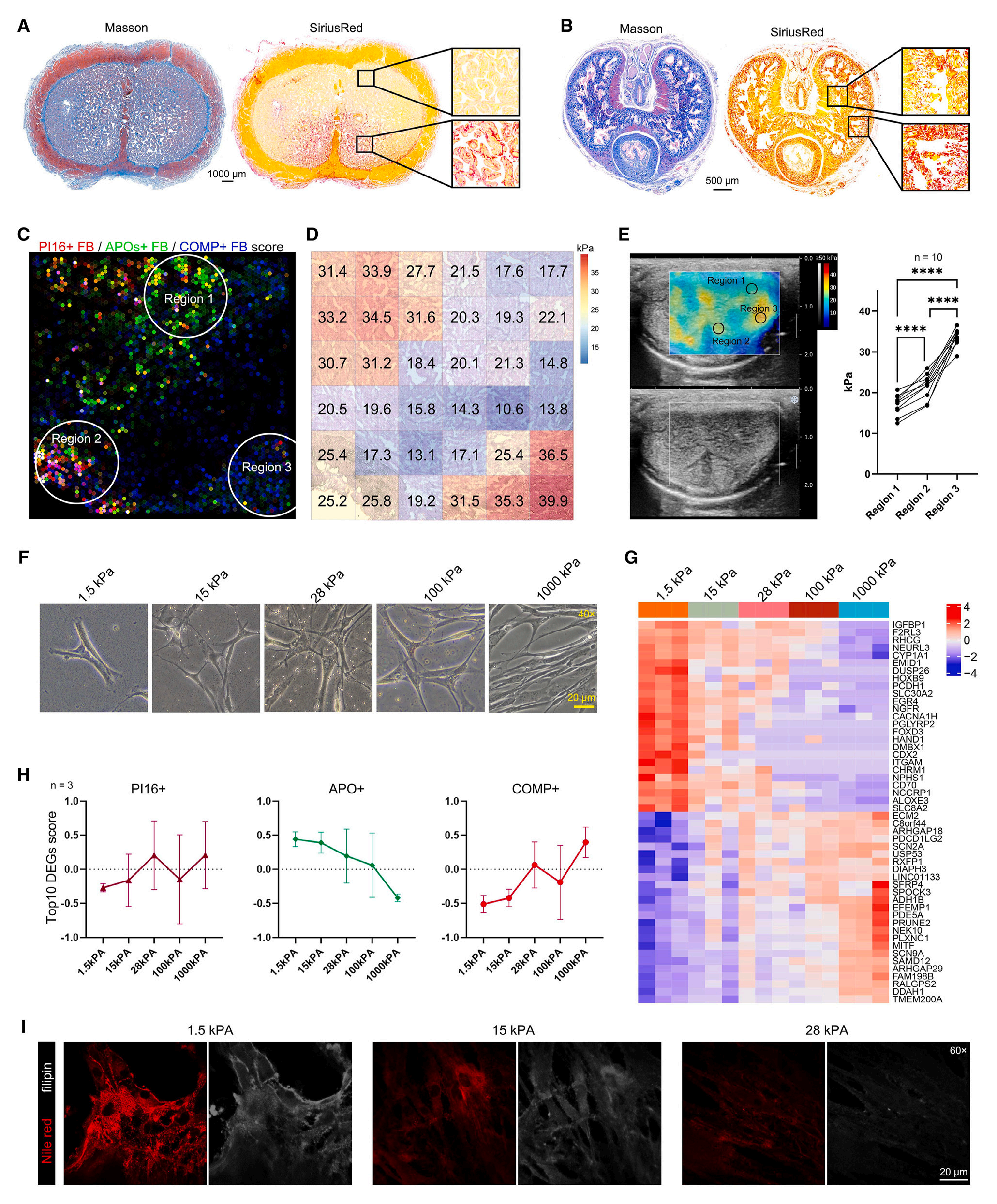
图7-局部机械力信号强度决定了这个区域内成纤维细胞的表型
10.人类和大鼠CC经历了相似的DMED病理变化
为了更好的认识DMED病理条件下,人类和大鼠CC微环境发生的变化是否与每种CC细胞类型中发生的分子水平变化相一致,研究者通过分析scRNA-seq数据,发现DMED大鼠模型和DMED患者都经历了显著的纤维化以及细胞组分丢失,但两者间的转录组差异显著。富集分析结果显示,仅DMED大鼠模型富集了“extracellular structure organization”;“actin cytoskeleton or organization”和“oxidative phosphorylation”仅在人类DMED患者SMC中发现,“regulation of MARK cascade”仅在DMED大鼠模型SMC中富集;“rhythmic process”仅在人类DMED患者FB中富集,“leukocyte activation”仅在DMED大鼠模型FB中富集。
空间转录组数据显示DMED大鼠模型阴茎中,除尿道以外的组织区域转录表达水平整体降低,而scRNA-seq数据显示DMED状态细胞的转录水平高于正常状态的细胞,这或许意味着,可能是由于单位面积上细胞数量降低,导致空间转录组数据出现转录水平降低的现象。

附录图13-通过scRNA-seq和空间转录组评估DMED条件下CC的病理变化
研究总结
本文绘制了人类和大鼠阴茎海绵体(CC)的生理、病理条件下的空间转录组图谱,结合多组学数据,系统比较了两个物种间的异同。本研究成果为人类、大鼠阴茎海绵体的分子解剖学,以及跨细胞类型和区域的信号网络提供了清晰洞见。同时在分子水平比较了人类和大鼠的阴茎海绵体异同,为理解动物模型的可用性提供基础数据,推动临床前实验和临床转化更高效、可靠。文中还阐述了CC中机械力信号的空间分布异质性,并确认ECM机械信号可以调节FB表型转变,提出靶向APO+ FB或可成为未来治疗DMED的有效手段。最后,研究者也明确指出了本文中研究的局限性,为后续研究指明了方向。
参考文献:
Yin et al, Molecular and spatial signatures of human and rat corpus cavernosum physiopathological processes at single-cell resolution. Cell Rep. 2024 Sep 24;43(9):114760. doi: 10.1016/j.celrep.2024.114760. Epub 2024 Sep 18.




 京公网安备 11011302003368号
京公网安备 11011302003368号