

研究背景
植物体细胞具备较高的发育可塑性,可通过体细胞胚胎发生过程,不经受精作用直接发育形成完整植株,该过程在作物无性繁殖、脱毒育种等农业生产中具有重要应用价值。已有研究表明,DNA 甲基化等表观遗传修饰参与体细胞胚胎发生过程,但其调控细胞全能性基因激活、介导体细胞重编程的具体分子机制尚不明确。
2025年5月,山东农业大学苏英华教授、张宪省教授团队联合德国研究人员,在Nature Communications发表题为“LEC2 induces somatic cell reprogramming through epigenetic activation of plant cell totipotency regulators”的研究成果,以拟南芥为材料,解析了转录因子 LEC2 通过 RNA 介导的 DNA 甲基化(RdDM)通路、CHH 甲基化识别复合体及染色质重塑调控,激活细胞全能性基因并诱导体细胞重编程的分子机制。
研究结果
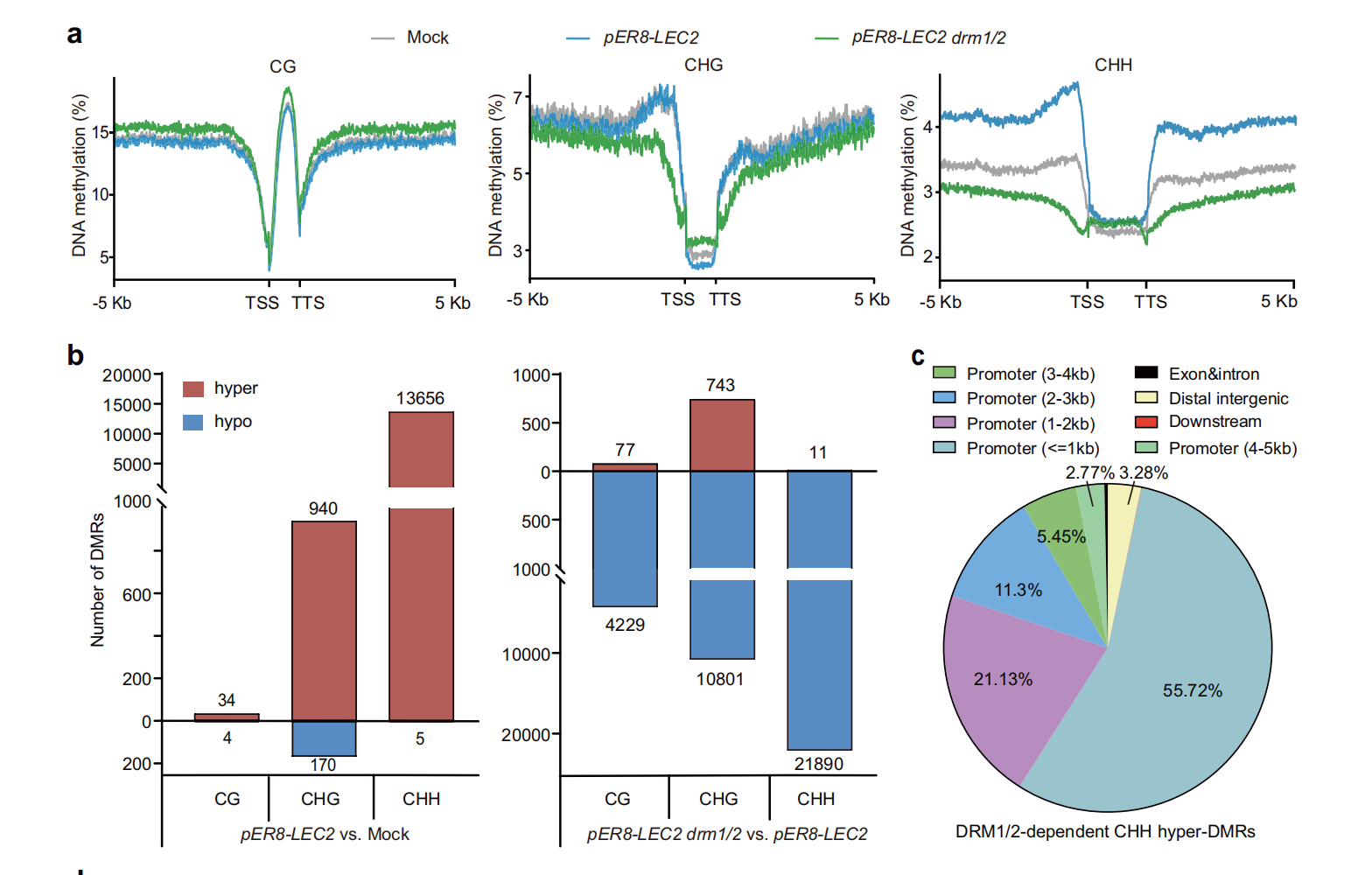
LEC2过表达通过激活DRM2/RdDM通路介导CHH高甲基化激活多能性调控基因过表达 LEC2 可显著上调 RdDM 通路核心基因表达,其中 DNA 甲基转移酶DRM2是 LEC2 的直接靶基因,LEC2 通过结合 DRM2 启动子区域的 RY 基序直接激活其转录。全基因组甲基化测序(WGBS)表明,LEC2 诱导后,基因组CHH 位点发生特异性高甲基化,CG 与 CHG 甲基化水平无显著变化,且 CHH 高甲基化区域主要富集于全能性调控基因的启动子区域。敲除 DRM1/2 会显著抑制 LEC2 诱导的体细胞胚胎发生,证明DRM2/RdDM 通路介导的 CHH 高甲基化是体细胞重编程的必要条件。
SUVH-SDJ-AHL复合体通过重塑染色质结构,将CHH甲基化信号转化为基因激活
CHH 高甲基化区域可被SUVH-SDJ 识别复合体结合,其中 SUVH1 受 LEC2 直接转录激活,SUVH1/3 与 SDJ1/2/3 形成的复合体可特异性结合 CHH 高甲基化位点。功能实验证实,suvh1/3、sdj1/2/3 突变体均显著抑制 LEC2 诱导的体细胞胚胎发生,且该复合体可与 LEC2 蛋白直接相互作用,作为共激活因子增强 LEC2 对靶基因的结合与转录激活能力。
为明确染色质状态在该过程中的作用,研究采用ATAC-seq(染色质可及性测序) 分析染色质开放程度。结果表明,LEC2 诱导后,LEC1、BBM、ABI3、FUS3 等全能性调控基因的染色质可及性显著升高;而 suvh1/3 突变体中,上述基因的染色质开放程度显著降低,说明 SUVH-SDJ 复合体是染色质开放的关键调控因子。进一步研究发现,SUVH-SDJ 复合体会招募AHL 家族染色质修饰蛋白,其中 AHL15 发挥主导作用。AHL 蛋白通过提升组蛋白 H3K9ac、H3K14ac 乙酰化水平,进一步促进染色质解凝缩,提升染色质可及性,进而保障全能性调控基因的高效转录。ahl15、ahl15/19 突变体均显著降低 LEC2 诱导的体细胞胚胎发生效率,且 ATAC-seq 结果显示,ahl15 突变会显著抑制全能性基因的染色质开放,验证了 AHL 蛋白在染色质重塑与体细胞重编程中的核心功能。
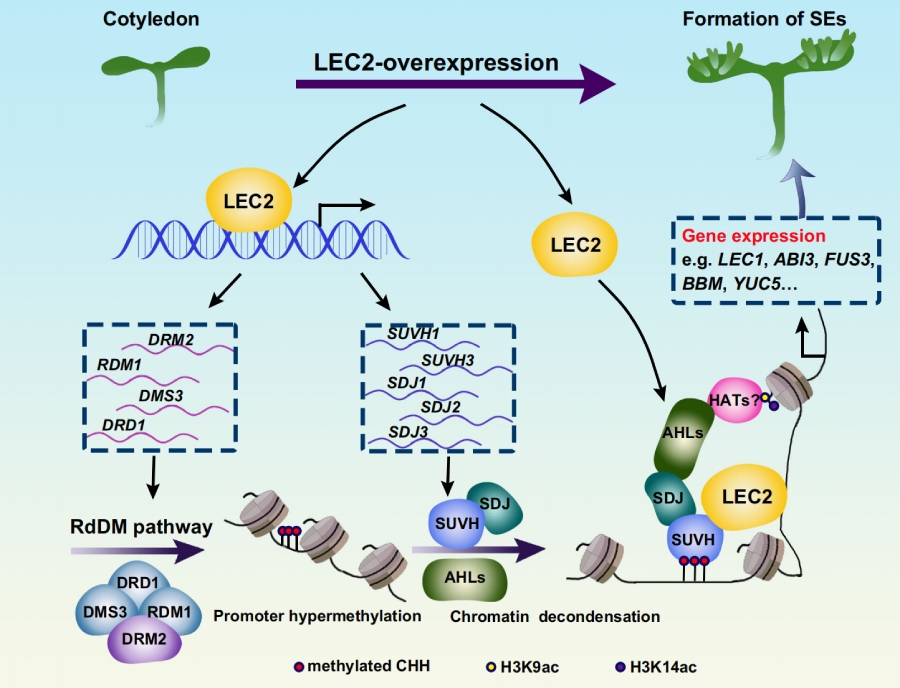
综上,本研究揭示了LEC2-DRM2-CHH 高甲基化 – SUVH-SDJ-AHL的表观遗传调控通路:LEC2 直接激活 RdDM 通路,在全能性基因启动子区域建立 CHH 高甲基化修饰;SUVH-SDJ 复合体识别该甲基化信号,并与 LEC2 协同作用;同时招募 AHL 蛋白提升组蛋白乙酰化水平、增加染色质可及性,最终激活全能性调控基因,完成体细胞向胚性细胞的重编程。
该研究系统解析了 DNA 甲基化、染色质重塑与植物细胞全能性激活的关联机制,完善了体细胞胚胎发生的表观遗传调控网络,为作物再生效率提升、育种技术优化提供了理论依据与分子靶点。



 京公网安备 11011302003368号
京公网安备 11011302003368号