
材料方法
结果
1. 基因组组装
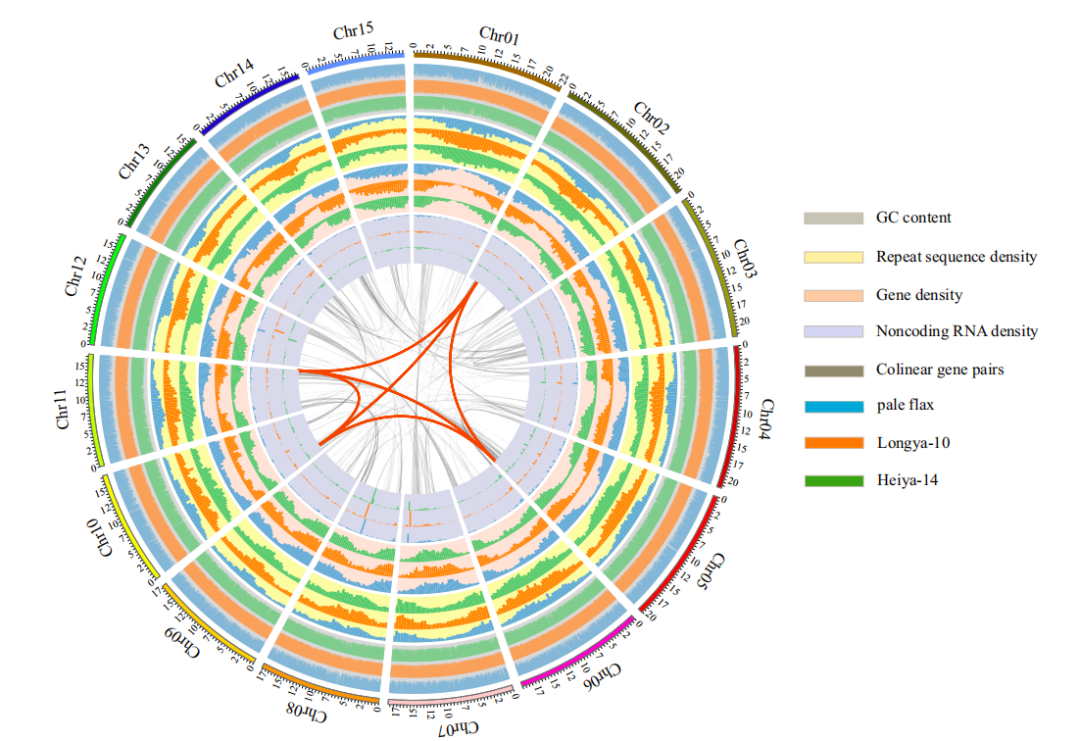
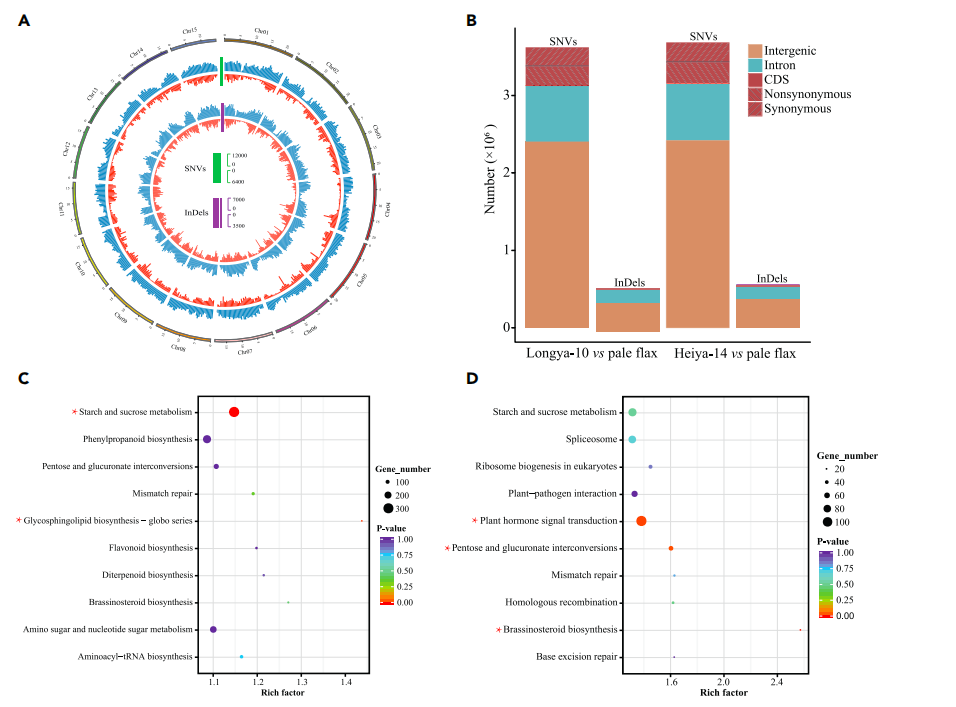
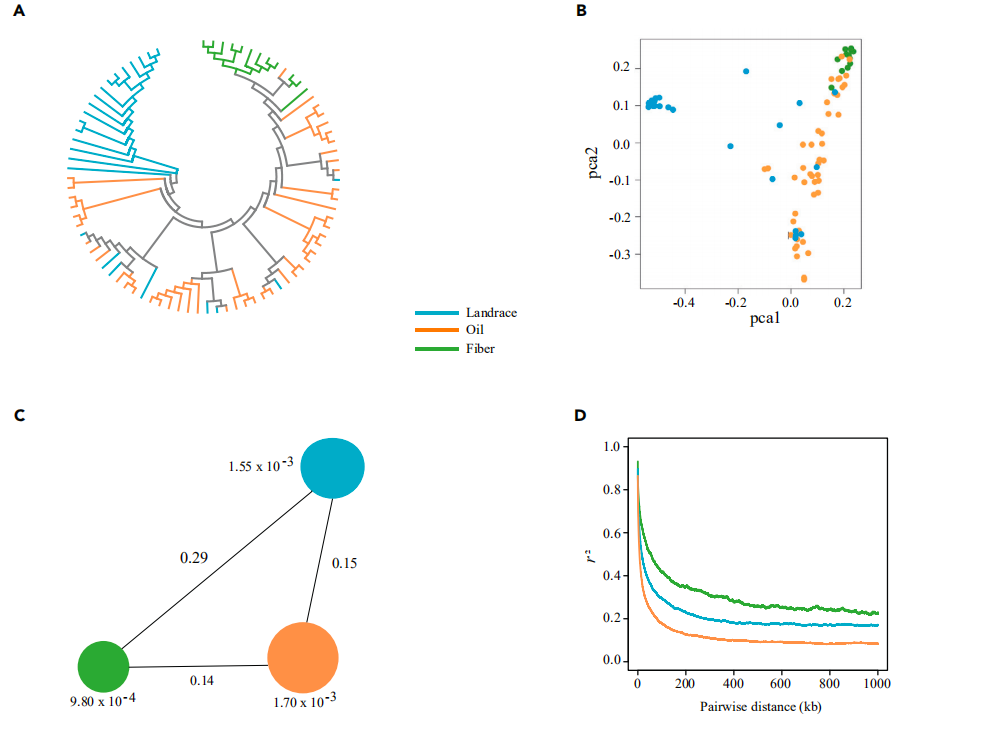
对油用亚麻品种Longya-10、纤维亚麻品种Heiya-14和pale flax分别测序得到68.2Gb,73.5Gb和 49.1Gb数据,组装出306.0Mb、303.7Mb和293.5 Mb基因组,其中contig N50/scaffold N50分别为131Kb/ 1,235Kb, 156Kb/700Kb和59Kb/384 Kb,后期利用HiC技术与遗传图谱辅助Longya-10基因组组装,将434 scaffolds组装至染色体水平,在每个基因组中预测到大约43,500个编码基因和2600-2800个非编码RNA;系统发育分析表明,栽培亚麻和白亚麻在大约232万年前发生了分化,自古被子植物的六倍化事件以来,亚麻发生了两次全基因组复制事件。

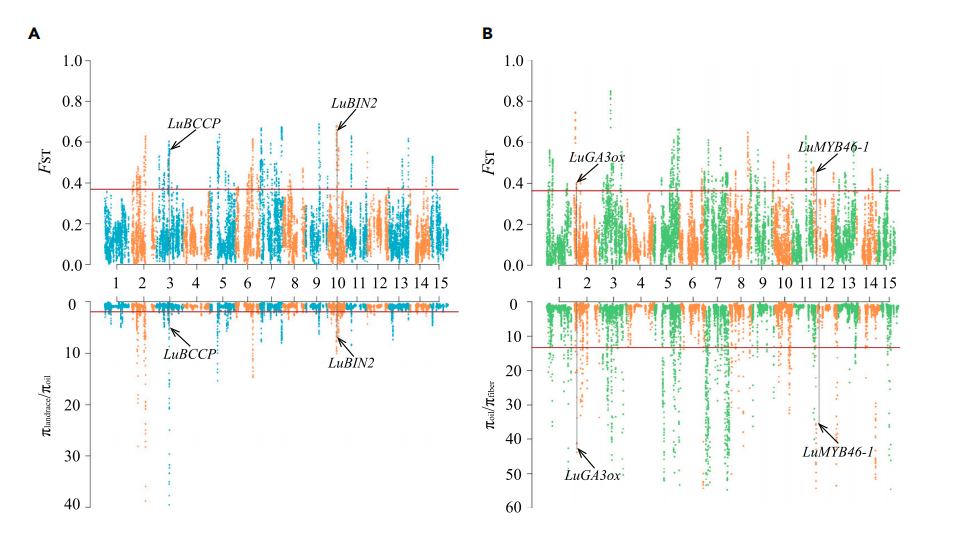
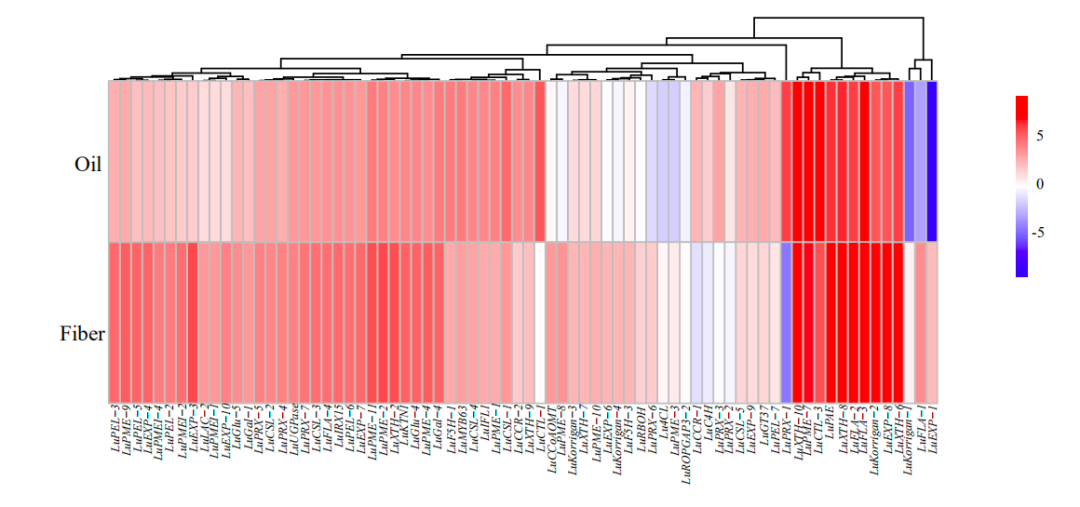
作物改良常常导致受选择的区域核酸多态性降低,将油用亚麻群体和纤维亚麻群体分别与地方种比较分析,油用亚麻中检测到108个选择清除区域(长度为15.5Mb,1,958个基因),纤维亚麻中有60个选择清除区域(长度为8.2Mb,1018个基因),27个选择清除区域重叠。其中α生物素羧基载体蛋白(LuBCCP)、脂氧合酶(LuLOX)、脂肪酰基ACP硫酯酶A(LuFatA)、脂质转移蛋白(LuLTP)和丙酮酸脱氢酶复合物E2组分(LuPDH-E2)等油脂相关基因以及种子大小相关基因LuBIN2和LuGW5在油用亚麻中受到选择。纤维亚麻中,次生细胞壁生物合成相关基因(LuMYB46-1、LuXTH和LuROPGAP3)和植物株高相关基因(LuGA3ox、LuGA20ox和LuGID)受到选择。
之前有报道纤维亚麻是从油用亚麻驯化而来,将油用亚麻与纤维亚麻进行比较分析,存在47个选择清除区域,其中50.9%与纤维亚麻相对于地方种的选择区域重合。这表明在纤维用亚麻改良过程中,这些区域持续受到很强的选择压力。大约一半的基因(426/867个基因)仅在油用亚麻和纤维亚麻比较中发现,包含内切-β-1,4-葡聚糖酶(LuKorrigan)、果胶甲酯酶(LuPME)和焦磷酸铜合成酶(LuCPS)等。作者将选择清除区域与之前报道的QTL/GWAS位点比较分析,发现2个硬脂酸相关的QTL与油用亚麻选择区域重叠,一个株高相关的GWAS位点与纤维亚麻选择区域重叠,另外3个油合成相关的QTL与纤维亚麻选择区域重叠,这说明亚麻在驯化过程中受到双重选择。
纤维是植物中一种特殊的细胞,具有加厚的次生细胞壁。AtMYB83/MYB46是拟南芥次生细胞壁生物合成的主要调控因子。研究发现,在亚麻、杨树、苹果、苜蓿和木薯中,MYB46/MYB83基因发生了物种特异性复制。8个已鉴定的LuMYB46/LuMYB83同源基因,4个在栽培种中高表达。此外,LuMYB46-1、LuMYB43-2和LuMYB83-1在栽培亚麻中发生了多种变异。与白亚麻相比,LuMYB83-1在两个栽培种中检测到21bp的插入,LuMYB46-1在亚麻改良期间受到了强烈的选择。LuMYB46-2在两个栽培种之间也有不同的插入/缺失变异。在亚麻中,四对MYB46/MYB83姊妹基因位于共线基因组区域,最近的一次分裂发生在最近一次WGD发生的时间,这意味着这次WGD事件导致了MYB46/MYB83基因在亚麻基因组中新的一次扩增。这些结果表明,在亚麻驯化和改良过程中,LuMYB46/LuMYB83基因的扩张和定向选择可能在重塑亚麻形态结构中起着至关重要的作用,qRT-PCR与RNA-Seq数据再次证明此结论。






 京公网安备 11011302003368号
京公网安备 11011302003368号