

2026年4月24日,花生功能基因组与种质资源创新实验室主任刘晓芹研究员团队联合何航团队和澳大利莫道克大学Rajeev K. Varshney院士团队在国际著名期刊《Nature Genetics》上发表了题为“Telomere-to-telomere genome assemblies and population resequencing of diploid and allotetraploid peanut varieties”的研究论文。该研究首次完成了2个野生祖先种(AA; BB) 及4个四倍体花生变种(var. hypogaea, var. hirsuta, var. fastigiata, var.vulgaris) 的T2T基因组构建工作。解决了现有基因组中存在较多gap和未组装到染色体水平的问题;完善了 var. hirsute 和var. fastigiate 主要栽培变种基因组信息空白的现状;为探究着丝粒区间的演化规律提供了基因组信息;通过521份代表性材料表型-基因型关联分析,挖掘解析了含油量、种子大小、脱水率等多个重要性状关键基因的分子调控网络。

花生是全球重要油料和经济作物,对保障世界粮油安全意义重大。花生起源于南美洲,在后哥伦布时期广泛传播,经过自然演变和人工驯化,产生了大量高产优质的地方品系和育成品种,这些品系、品种为花生的改良提供了宝贵的遗传资源。但受限于基因组高度复杂、为异源四倍体等特点,花生基因组学研究明显滞后于其他作物,严重制约了花生遗传资源利用与育种进程。因此,构建花生 T2T 基因组、高效挖掘基因资源及解析关键农业性状基因,是培育高产、高油、宜机收、脱水快等优良新品种、保障粮油安全的关键途径与重要方向。
1.花生T2T基因组构建及评估
本研究选用了 2 个野生二倍体祖先种和 4 个四倍体花生变种,,利用HiFi、ONT 以及 Hi-C 数据开展基因组组装与挂载工作(图1)。组装和评估后Contig N50 平均值达到 85.94Mb,基因组大小在 1.18Gb 到 2.63Gb 之间;基因组 BUSCO 平均值为 98.67%,LAI 为 21.76,QV 为 47.52;二代数据平均比对率 99.76%,转录组比对率 92.93%,注释出 34,406 至 75,143 个蛋白编码基因,平均基因功能注释率达 98.75%。因此获得连续性好、完整度高,以及高质量的基因注释的基因组。
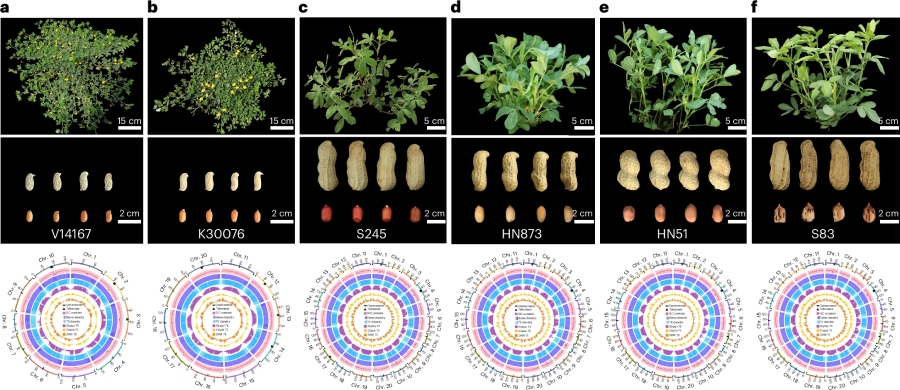
图1 花生表型及基因组组成特征。
2.亚基因组间不对称进化
通过对亚基因组间转座子(TE)的比较分析发现,A (t) 和 B (t) 亚基因组中分别含有约 605.50 Mb 和 865.57 Mb 的 Gypsy 元件,其可能是造成A (t) 亚基因组小于 B (t) 亚基因组的主要原因。进一步分析显示,Gypsy 在 A (t) 亚基因组中发生过一次插入事件,而在 B (t) 亚基因组中发生过两次插入事件,且这些事件在二倍体祖先种中就已出现(图2)。基于全长 Althila-TE 构建的系统发育树表明,A、B 基因组分化后,Althila-TE 呈现出独立演化的特征。
对着丝粒区域的分析发现,At 亚基因组的着丝粒平均长度为 46.11 Mb,与 A 基因组的 44.70 Mb 基本一致;而 Bt 亚基因组着丝粒仅 25.70 Mb,明显小于 B 基因组的 43.30 Mb。进一步分析发现造成该差异的主要原因是重复单元数目减少造成的。我们推测,这一差异主要是为了保证四倍体中同源染色体之间的正常交换而产生的适应性进化。
此外,对着丝粒区域 CentO 重复单元的进化树分析显示,其在 A (t) 和 B (t) 亚基因组中独立进化,而 CRMs 则表现为协同进化。同时对比发现,相较于 B (t) 亚基因组,A (t) 亚基因组在近期发生了更多的插入事件;与二倍体祖先相比,四倍体栽培种的着丝粒区间经历了明显的重塑事件。这说明着丝粒在快速演化的同时,也保持了较高的保守性。
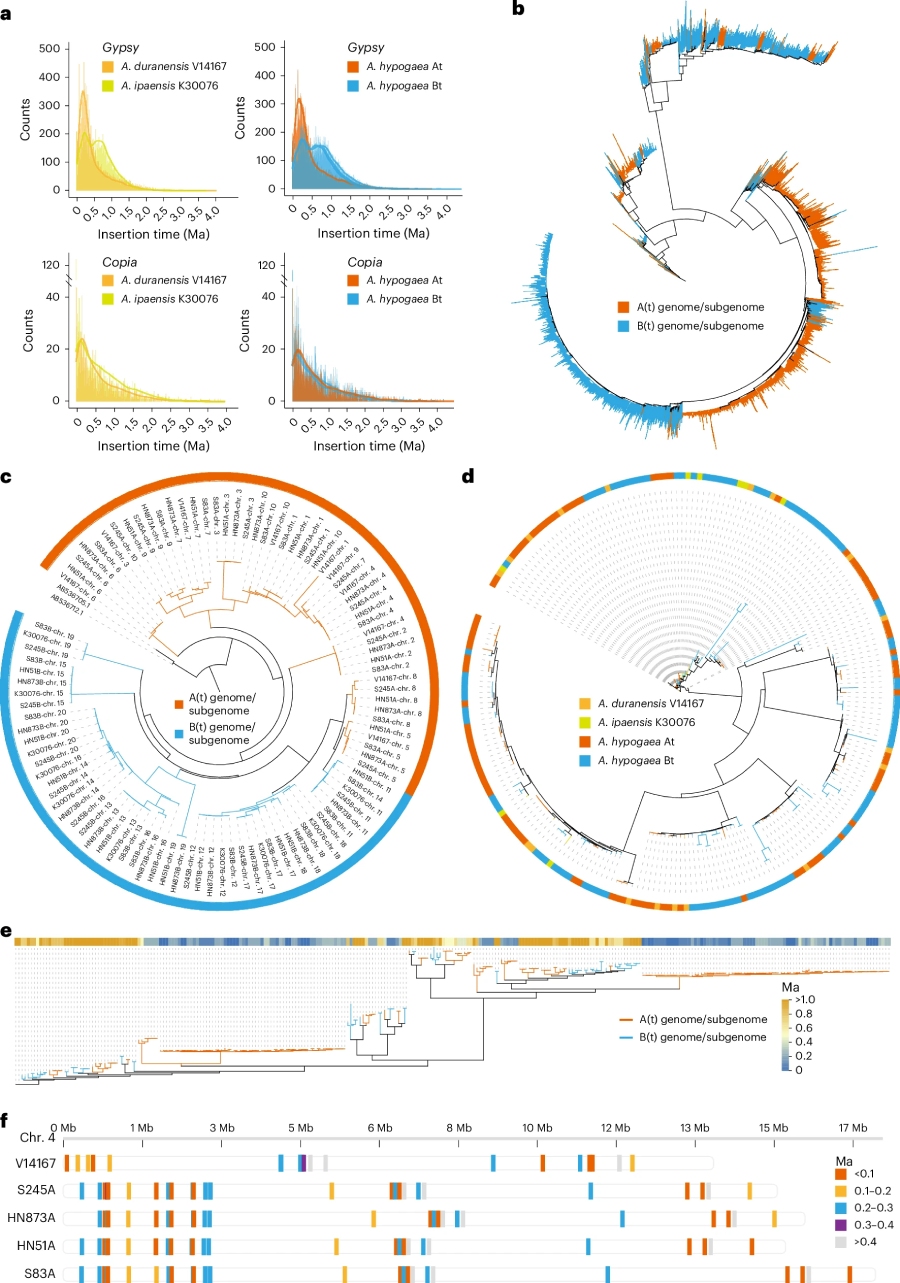
图2.亚基因组进化分析
3.变种间结构变异
基因组比较分析发现,相比于V14167_vs_At,K30076_vs_Bt表现出非常高的共线性,进一步提供了其作为四倍体祖先种的证据。进一步比较分析了四倍体变种特异SV及SV-genes,证实了SVs在驯化过程中塑造花生遗传分化中的重要作用(图3)。另外,在Chr14上鉴定到13.23 Mb 的Indel;利用Illumina,HiFi及Hi-C分析证实了该SV的可靠性。该SV中的109个基因主要富集在脂质代谢,环境响应,器官发育等term;进一步分析发现,来自同一亚种的var. hirsute (161/161存在插入) 和 var. hypogaea (33/161存在插入)变种之间存在着明显株型差异,同源比对发现该变异内基因的同源基因有OsWOS3A/3B,OsLAC 等与器官发育相关的关键基因。暗示在驯化过程中,这些SV-gene在塑造同一亚种间不同变种之间的株型差异发挥了重要作用。
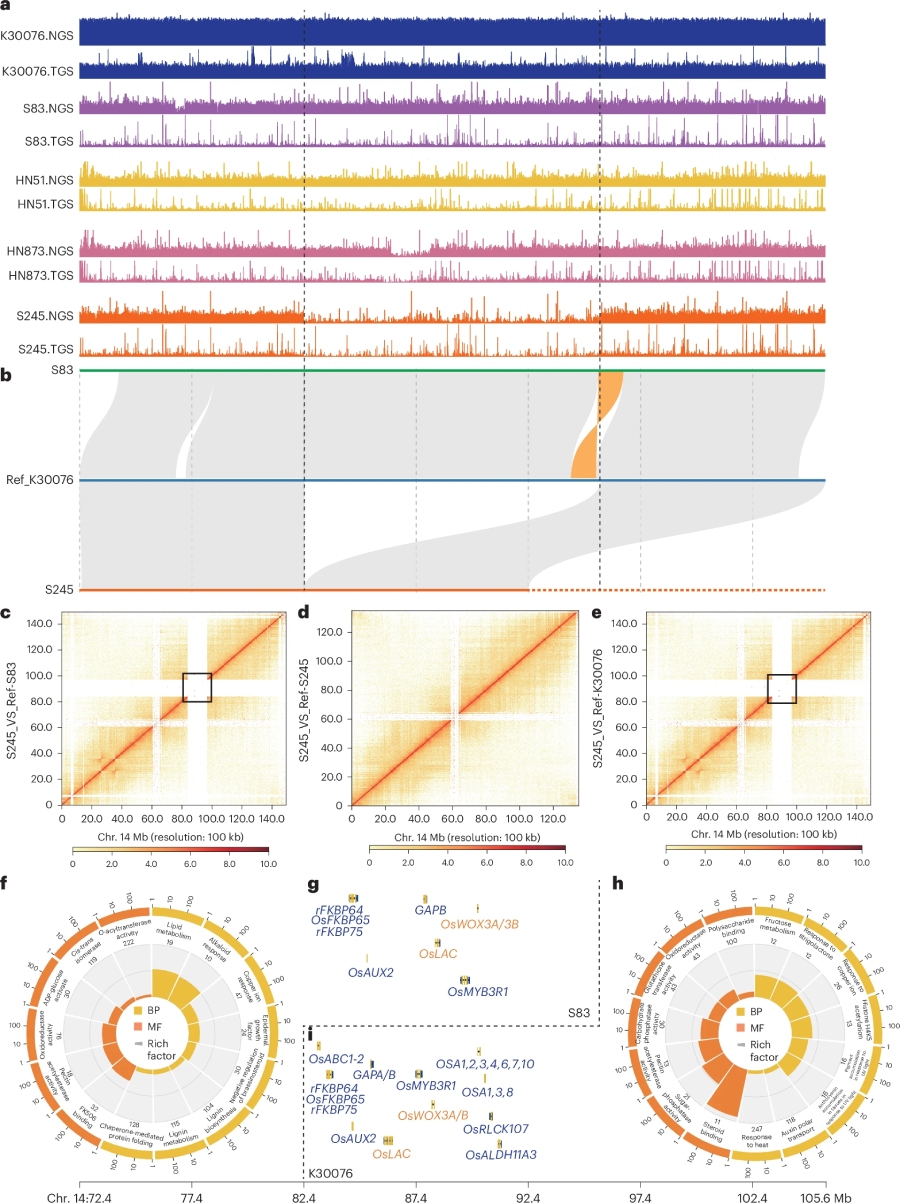
图3.结构变异分析
4.群体遗传分析
群体分析选取了 521 份代表性的花生材料,以 S245 T2T 基因组作为参考,鉴定出 101Mb 的高质量 SNP 变异用于群体分析。结果显示(图4),除了野生群体之外,这些材料能明显分成两个亚群和四个变种群,说明不同变种之间存在明显的遗传分化。其中比较有意思的是 Adm 群,通过材料系谱分析发现,该群体中的大部分材料,都有利用野生材料进行改良的过程,如远杂 9102等。另外,选择和渗入分析表明, At 和 Bt 亚基因组之间存在着明显的不对称进化结果。
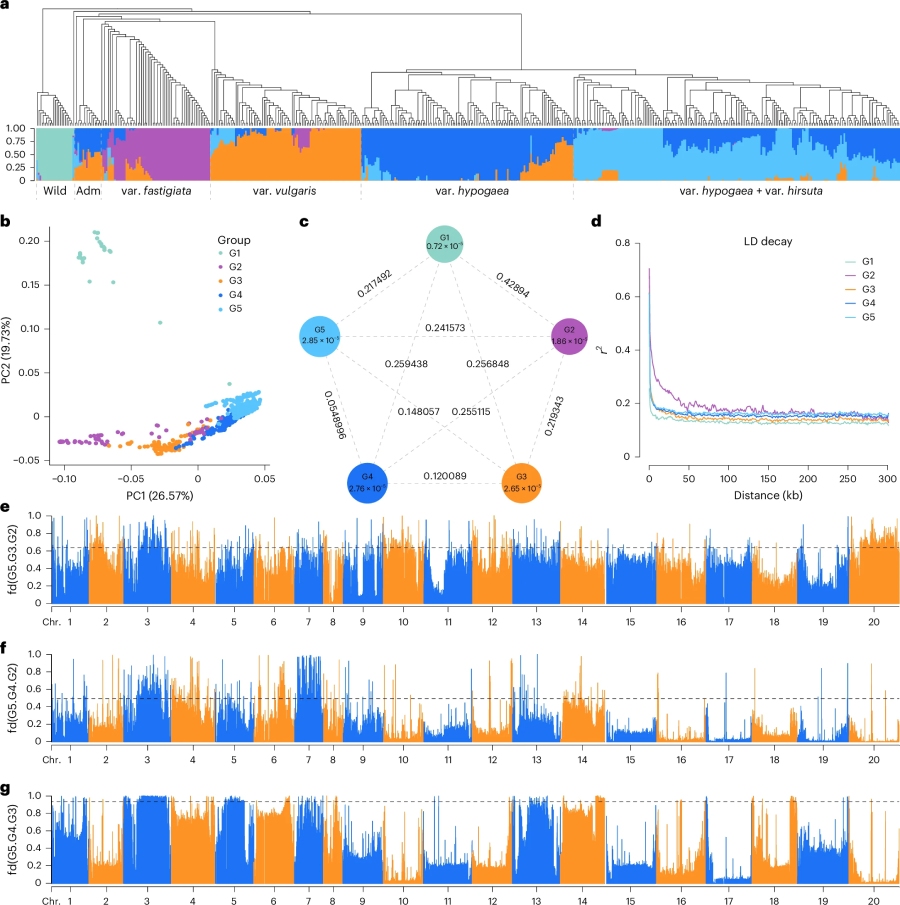
图4.群体遗传分析
5.含油量和种子重量Causal基因及单倍型鉴定
GWAS及Fst 在chr08上鉴定到含油量性状显著相关信号;进一步鉴定到关键候选基因AhWRI1; 在启动子及第一个外显子上存在单倍型差异,并且与表型显著相关;实验结果表明:启动子区域突变(A>C)是通过影响AhFUS3与 AhWRI1基因的结合,进而影响其调控的下游基因AhACP1和AhKCS1;导致不同花生品种油含量差异(图5)。同时我们在chr16上鉴定到千粒重性状显著相关信号;进一步鉴定到关键候选基因AhGSA1; 基因结构分析发现,在启动子区间鉴定到碱基插入(AT > ATT)引起单倍型变化,且与表型显著相关;实验结果表明,Hap2 启动子活性显著高于Hap1,影响AhGSA1 基因的表达,进而引起千粒重大小差异。
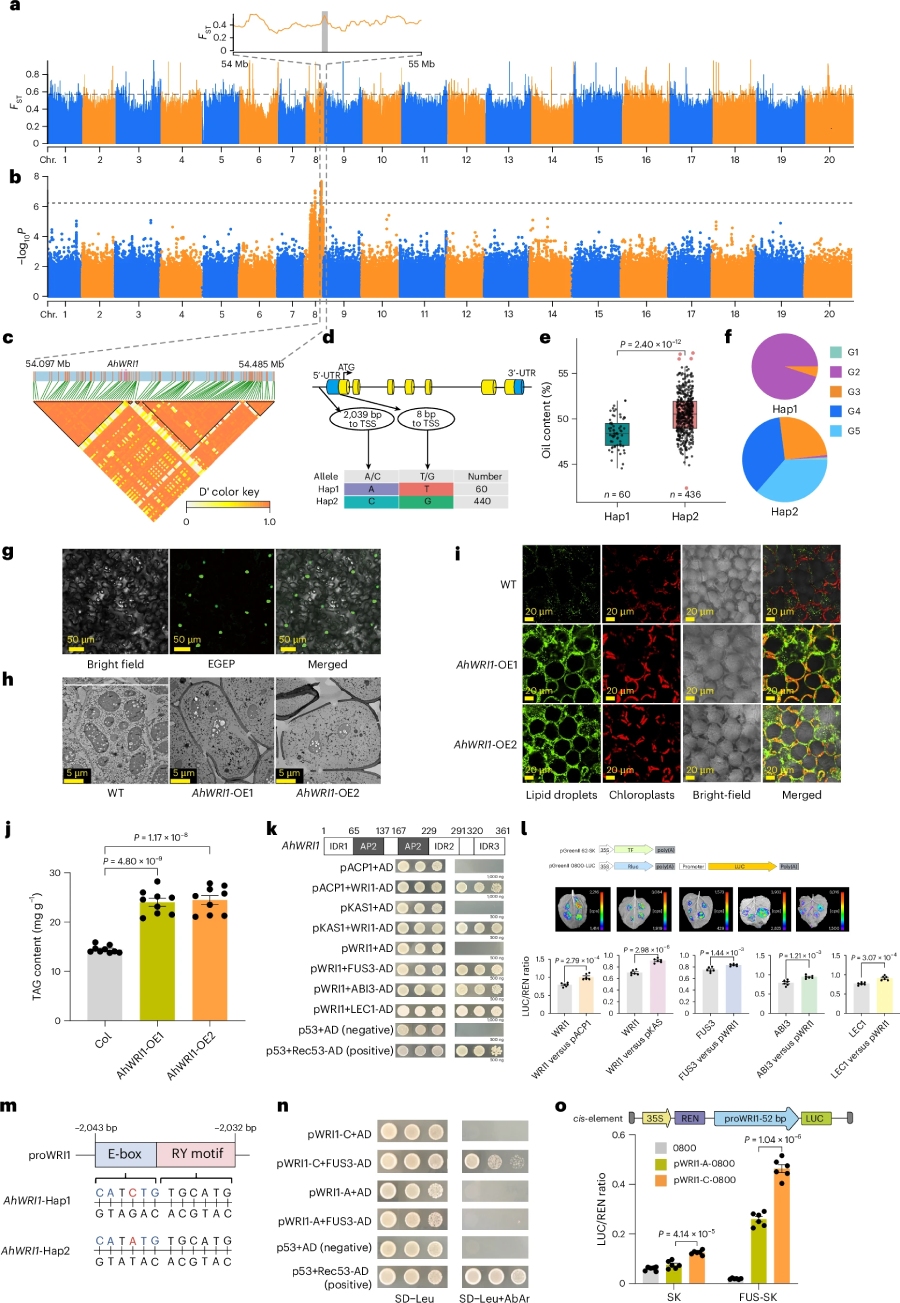
图5.油含量基因挖掘与鉴定
6.脂质和花青素动态变化图谱构建
为探究花生种子发育机制,我们对 S83 和 S245 两个品系的五个发育时期开展转录组与代谢组联合分析。研究共检测到 497 种脂质代谢物,其中 S83 的脂质含量显著高于 S245。经 WGCNA 分析,多个脂质代谢相关的酶基因在 S83 中表达量更高。编码 MYB 转录因子的 AhRt2 基因表达差异可能是调控S83 和 S245颜色分化的关键因素(图6)。本研究揭示了调控花生种子发育与种皮色泽的特异代谢及合成机制,为解析相关分子机理提供了重要依据。
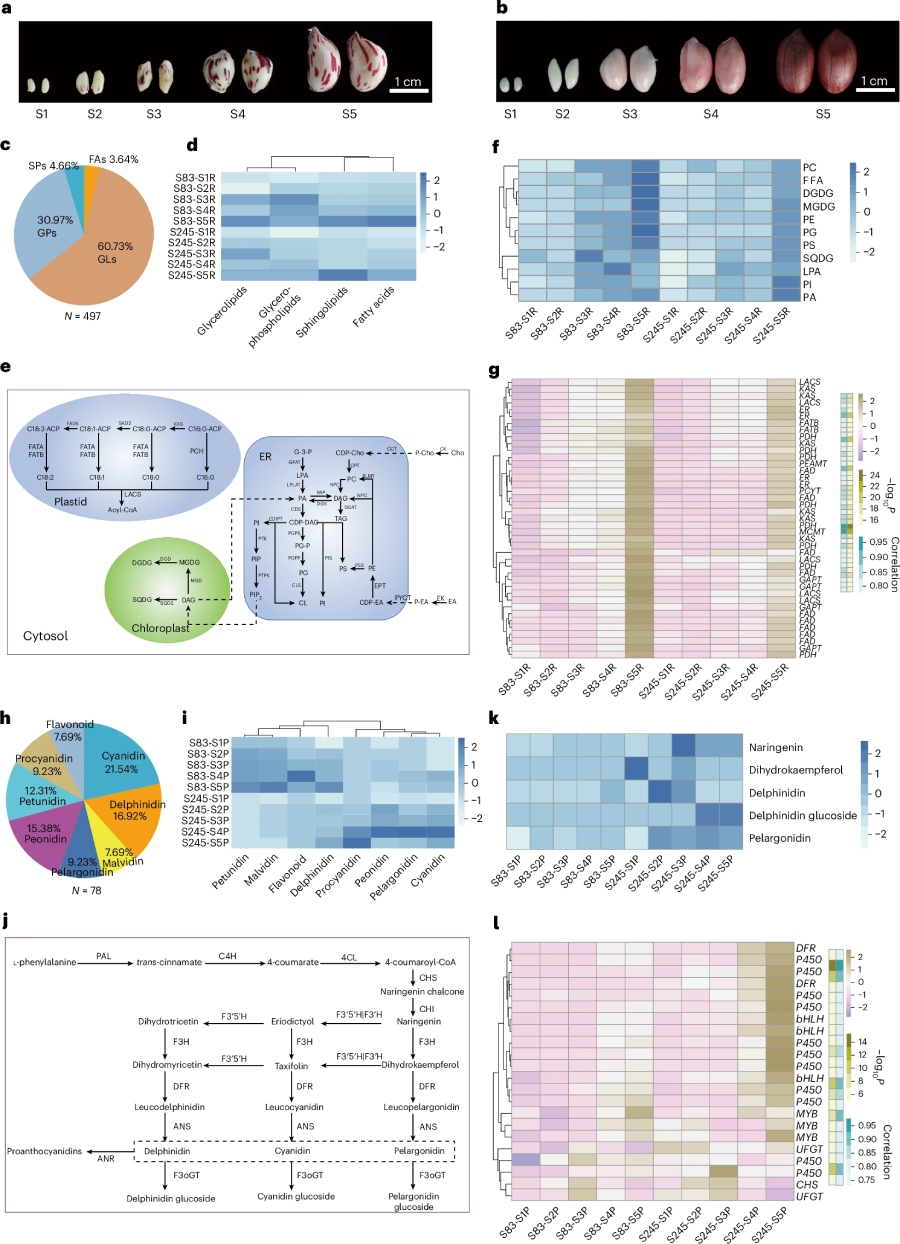
图6.脂质和花青素代谢通路解析
北京大学现代农业研究院花生功能基因组与种质资源创新实验室主任刘晓芹研究员,多组学实验室何航研究员和澳大利亚莫道克大学Rajeev K. Varshney院士为共同通讯作者。北京大学现代农业研究院卞建新副研究员,北京大学张宜林博士,北京大学现代农业研究院科研助理丁帅,莫道克大学联培生郭昊松为论文共同第一作者;李奎副研究员,科研助理管玉、李季华、崔媛媛助理研究员、莫道克大学Vanika Garg 和Annapurna Chitikineni,科研助理孟庆静、周国靓、吕源、李甜雨为本研究做出了重要贡献。北京大学现代农业研究院邓兴旺院士、广西省农科院贺梁琼研究员和唐荣华研究员、山东省农科院王兴军研究员和赵传志研究员、浙江大学张亮生教授以及北京大学现代农业研究院高性能计算中心为本研究提供了重要支持。
该研究得到山东省重点研发计划农业良种工程、山东省泰山学者项目、山东省自然科学基金和潍坊现代农业山东省实验室等基金的资助。
引用文献:
Bian, J., Zhang, Y., Ding, S. et al. Telomere-to-telomere genome assemblies and population resequencing of diploid and allotetraploid peanut varieties. Nat Genet (2026).
DOI:10.1038/s41588-026-02577-z




 京公网安备 11011302003368号
京公网安备 11011302003368号