

2026年4月,沈阳农业大学植物保护学院纪明山教授研究团队,在一区Top期刊The Plant Journal 发表了题为“Genomic insights into herbicide resistance evolution in allopolyploid Monochoria korsakowii ”的研究论文。

该研究首次获得了雨久花染色体水平基因组,并分析了其基因组进化特征,对参与苄嘧磺隆(BSM)代谢的关键代谢酶编码基因进行了分析并验证了其抗性机制,这些结果阐明了雨久花适应的潜在基因组机制,为制定基于基因组的精准杂草防控策略奠定了关键基础。百迈客生物为该研究提供了基因组测序、分析等服务。
研究背景
雨久花(Monochoria korsakowii)是水稻田中常见的阔叶杂草,严重危害水稻生产,田间种群已普遍对ALS类除草剂苄嘧磺隆(BSM)产生抗性,传统防除方案失效;该物种也被证实具有金属耐受性。然而,目前雨久花的基因组信息仍十分匮乏,阻碍了对其除草剂抗性和金属耐受性遗传机制的全面解析。
研究结果
雨久花染色体水平基因组
该研究证实雨久花为四倍体,染色体数目为2n=4x=56,结合流式细胞术、K-mer分析明确了其基因组大小与重复序列占比。研究利用高通量数据完成基因组组装,组装大小1.87 Gb,BUSCO评分、测序读长比对率等指标均证明组装质量优异;经Hi-C挂载将绝大部分序列锚定并定向到28条假染色体,染色质互作热图、荧光原位杂交结果也进一步验证了组装效果与该物种的四倍体属性,其基因组特征与同属二倍体高亭雨久花存在明显差异。
研究依据15-mer序列将28条假染色体划分为A、B两个亚基因组,并统计了两个亚基因组的序列长度、染色体数量及支架指标。通过多方法联合注释,共鉴定出67,496个高完整度蛋白质编码基因,基因功能注释覆盖率较高,同时识别出丰富的非编码RNA;该基因组重复序列占比60.74%,其中长末端重复逆转录转座子为优势重复元件。
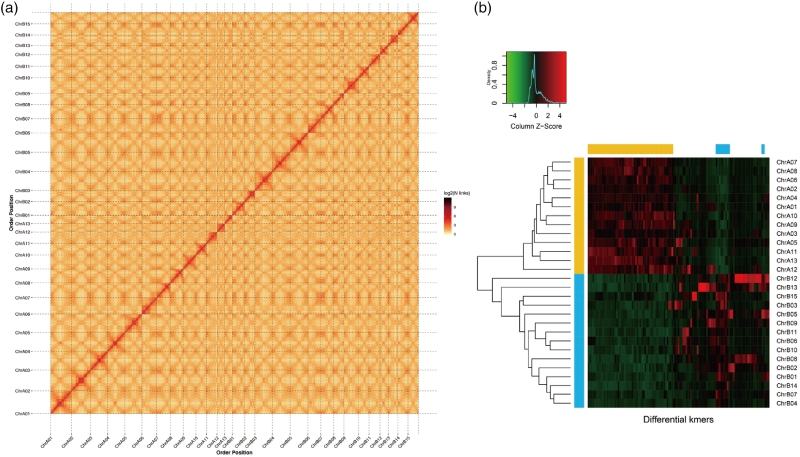
图1 异源四倍体雨久花基因组
基因家族进化与分化时间估算
该研究从雨久花中鉴定出359个物种特异性基因家族,共计1462个基因,两个亚基因组的基因家族呈现出截然不同的功能富集特征。亚基因组A的特有家族主要富集于核酸酶活性、转移酶活性以及抗氧化解毒、硒化合物代谢等通路;亚基因组B的特有家族则集中在能量代谢、植物生长发育调控相关通路。此外,两个亚基因组分别存在数量可观的扩张基因家族,功能各有分工,部分基因家族还表现出亚基因组间收缩与扩张的差异,证明亚基因组之间存在明显的功能分化。
基于463个单拷贝直系同源基因构建系统发育树并估算分化时间,结果表明该物种的两个亚基因组拥有不同祖先起源。亚基因组A与高亭雨久花互为姐妹支系,A、B 两个亚基因组约在881万年前发生分化,这一时间早于亚基因组A与高亭雨久花祖先的分化时间(665万年前)。
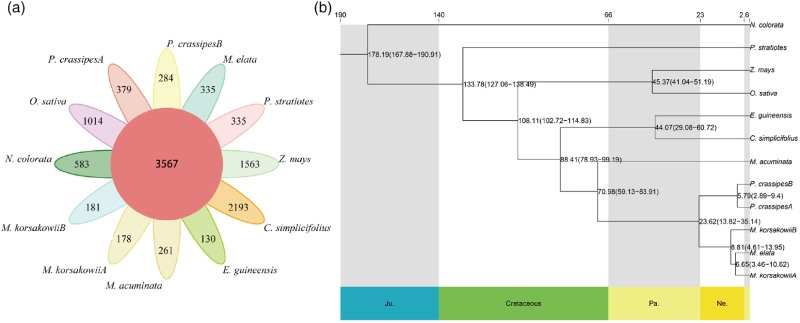
图2 基因家族进化与系统发育分析
全基因组复制及重复基因功能富集分析
同义替换率(Ks)分析表明雨久花经历过多次全基因组复制,证实其为异源多倍体,物种间遗传分化规律也与系统发育结论相符。研究在该物种基因组中鉴定出92,532对重复基因,并分为五类复制类型。功能富集结果显示,全基因组复制来源的基因主要参与激素相关应答过程;串联重复基因富集于除草剂解毒相关通路,可依靠基因剂量扩增帮助植株快速适应环境;其余几类重复基因则集中在碳水化合物结合、ADP结合等分子功能上。
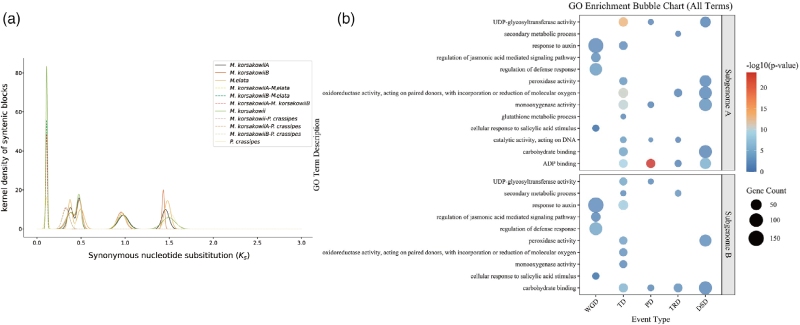
图3 多倍体雨久花的亚基因组进化
雨久花科祖先核型重建及核型进化
通过对雨久花科三个物种的基因组和亚基因组进行共线性分析,重构了它们核型的进化历史。在一次古老的全基因组复制以及随后的染色体重排之后,祖先核型最终形成了高亭雨久花(14条染色体)、雨久花亚基因组A(13条染色体)和雨久花亚基因组B(15条染色体)各自独特的核型。这一重构的核型框架为理解基因家族的组织与进化提供了基因组背景,其中包括潜在参与抗除草剂机制的基因家族。
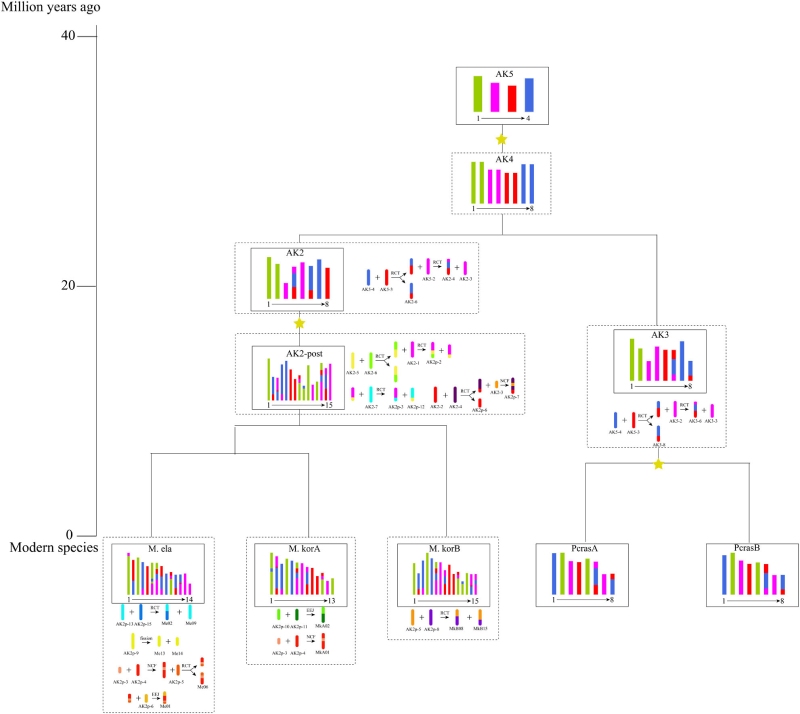
图4 三种雨久花科植物核型进化的重建
雨久花的基因组分化偏倚及亚基因组优势
该物种的两个亚基因组呈现不对称进化特征:B亚基因组基因数量更多,而A亚基因组基因丢失程度更高、保守基因缺失比例更大,进化速率相对更快,二者转座元件分布则无明显差异。共线性分析结果也进一步证实了A亚基因组基因丢失更显著的特点。
基因表达分析显示,43.6–54.6%的单拷贝同源基因存在亚基因组偏向性表达,但整体未形成亚基因组表达显性,同时存在两类分别持续偏向A、B亚基因组表达的基因。功能富集表明,两类偏向表达基因参与的通路出现分化,分别关联胁迫耐受、光合稳态、活性氧清除等生理过程,部分代谢通路由两个亚基因组协同完成。选择压力分析显示,发生偏向表达的基因拷贝受到的纯化选择作用更强。
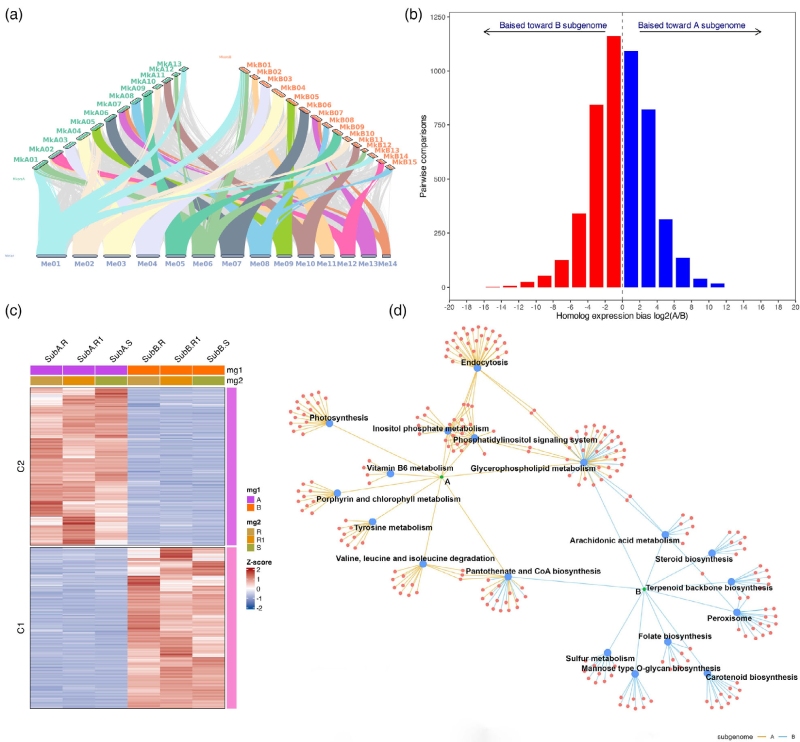
图5 多倍体亚基因组分化驱动雨久花产生功能特化
雨久花苄嘧磺隆(BSM)抗性候选基因筛选及表达验证
研究利用马拉硫磷(细胞色素P450抑制剂)与NBD-Cl(谷胱甘肽S-转移酶GST抑制剂)开展药剂预处理试验,结果显示两种抑制剂均可削弱抗性种群对苄嘧磺隆的耐药性,其中NBD-Cl作用效果远为显著。酶活性动态检测表明,除草剂处理后抗性种群的P450与GST 活性均高于敏感种群,且GST活性的提升幅度、作用持续时间均明显更强,证实GST介导的解毒通路是该物种耐受苄嘧磺隆的核心途径。
本研究共鉴定出77个GST基因,明确了其系统发育关系、染色体分布及保守基序特征。通过转录组分析筛选出3,474个差异表达基因,进一步得到7个在抗性种群中显著上调的GST 基因,并锁定3个关键候选基因,RT-qPCR验证了多数候选基因在多个抗性株系中稳定高表达。序列分析发现 GST基因启动子存在大量非生物胁迫相关顺式元件;构建基因-转录因子互作网络后筛选出一个核心转录因子,该因子在抗性种群中表达上调,推测其可调控GST基因表达,该推测仍需后续实验验证。
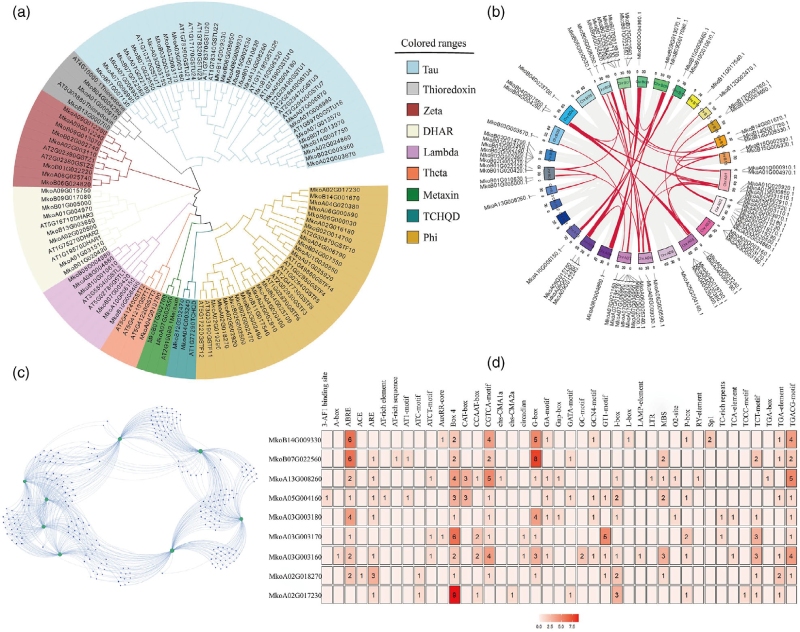
图6 系统发育、共线性、共表达及启动子分析解析雨久花GST基因家族
GST对BSM的动力学特性及代谢能力
研究将3个候选GST基因在大肠杆菌中异源表达并完成蛋白纯化,SDS-PAGE检测证实所得重组蛋白纯度高、分子量与预测结果一致。以CDNB为底物进行酶动力学分析,发现MkoA03G003170的催化效率与底物亲和力最佳,其余两种重组蛋白的催化活性和底物亲和力相对偏弱。体外代谢实验结合HPLC检测表明,三种蛋白均能降解苄嘧磺隆(BSM),其中MkoA03G003170对苄嘧磺隆的降解能力最为突出。
MkoA03G003170蛋白与BSM的虚拟分子对接
综合酶动力学与代谢实验结果可知,MkoA03G003170是降解苄嘧磺隆活性最优的重组蛋白,系统发育分析证实该蛋白属tau类谷胱甘肽S-转移酶家族。研究通过同源建模解析其三维结构,并完成与苄嘧磺隆的分子对接,明确了二者的结合模式、结合位点氨基酸组成、结合能及解离常数。亚细胞定位实验显示,该蛋白在本氏烟草中同时分布于细胞核与细胞质,推测这种定位特征使其具备多样的生物学功能。
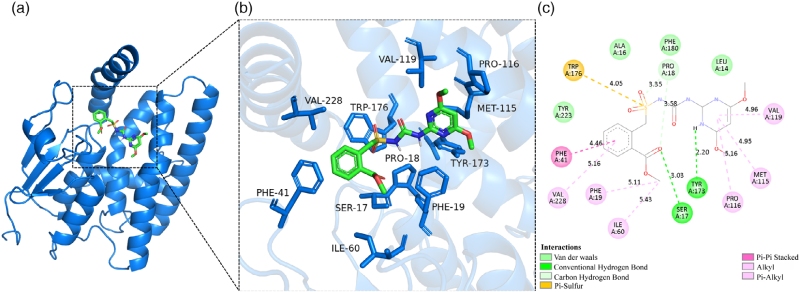
图7 雨久花MkoA03G003170蛋白与苄嘧磺隆的分子对接
研究总结
该研究首次完成异源四倍体雨久花染色体水平基因组组装,发现该物种两个亚基因组不存在整体显性,而是形成功能特化,分别富集不同代谢通路;串联重复基因与除草剂抗药性演化密切相关,并鉴定出介导苄嘧磺隆抗性的关键GST基因MkoA03G003170。该研究证实多倍体诱导的基因组可塑性是抗药性形成的潜在因素,为理解杂草适应性及制定可持续管理策略提供了基础基因组资源。




 京公网安备 11011302003368号
京公网安备 11011302003368号