

英文标题:Integrated Analysis of Metabolome and Transcriptome Reveals Insights forCold Tolerance in Rapeseed (Brassica napus L.)
发表期刊:Frontiers in Plant Science
发表时间:2021-10-8
影响因子:5.753
发表单位:中国农业科学院农业部油料作物生物学与遗传改良重点实验室,农业部油料作物研究所
研究背景
油菜(Brassica napus L.)是世界上重要的油料作物。低温胁迫(CS)等多种非生物胁迫对其生产力有显著影响,严重限制油菜的生长和产量,从而造成农业经济损失和生产风险,提高耐 CS 成为农业调查和作物改良的一个重要领域。耐寒性是作物抵御低温的基本特性,尤其是越冬作物。因此,开发耐冷油菜品种以保证在这种不利条件下的产量是当务之急。
目前,基因组学、转录组学、蛋白质组学、代谢组学等高通量组学方法已被植物研究者广泛应用于研究不同的非生物胁迫,加深了对不同生物途径的理解。本研究旨在通过结合转录组和代谢组分析,确定油菜品种的胁迫响应基因、代谢产物和代谢途径,以了解耐寒(C18)和耐寒(C6)油菜品种的低温胁迫响应和耐寒机制。本研究为油菜的生长机制提供了新的见解,研究了油菜抗逆性的影响因素,以促进油菜抗逆性品种的选育。
材料方法
材料:2 个油菜品种,耐寒品种 C18(CT-C18)和冷敏感品种 C6 (CS-C6),分别冷处理的 0、1、7 天后进行取样,每组样本设置三个生物学重复
组学方法:RNA-seq、LC-MS/MS
主要结果
一、冷胁迫下 C6 和 C18 品种的生理响应及代谢谱分析
表型结果表明,C6 在冷处理 7 天时表现出比 C18 严重的萎蔫和褪绿。CT-C18 在第 1天和第 7 天的 SOD、POD 等抗氧化酶活性较高,C18 具有更好的抗氧化能力。并且 C18的可溶性糖、可溶性蛋白和脯氨酸含量都有显著升高。以上的现象都表明 C18 可通过增加CS 的抗氧化防御酶活性和渗透保护剂含量,降低氧化应激,是比 C6 更好的耐寒品种。
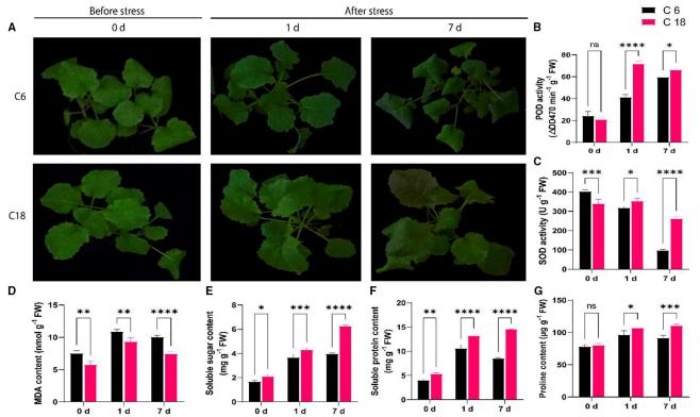
耐寒(C18)和冷敏感(C6)油菜品种在 0 d (CK)、胁迫 1d 和 7d 对冷胁迫(4℃)的生理指标
为了构建一个系统的代谢变化概况,对冷敏感(CS- c6)和耐寒(CT-C18)品种进行了非靶向代谢组分析。共检测到代谢物 3,368 种;其中,626 种是已知的代谢物。为了更好地了解CS 对代谢的影响,建立了无监督层次聚类,表明 CS 早期(0 1 天)的代谢数据与应激后期(7天)明显分离。PCA 的 PC1 显示出不同时间点之间的分离,上述结果表明,两品种间的代谢反应差异可能是其对冷胁迫耐受性不同的基础。
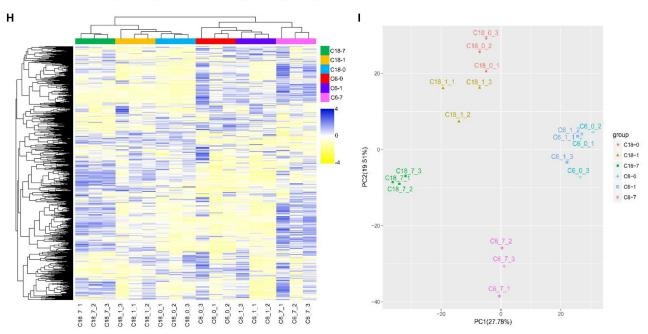
非靶向代谢组的无监督层次聚类分析和 PCA 分析
二、两个油菜品种间冷反应的研究
为了鉴定冷胁迫条件下油菜籽的差异代谢物,作者比较了各时间点对照条件下油菜籽的代谢产物数量和胁迫水平,C6 样本的 0、1、7 处理组中分别发现了 156、357、358 个差异基因,C18 样本的 0、1、7 处理组中分别发现了 182、755、469 个差异代谢物。而同一处理时间内的两个品种间的差异代谢物分别为 429、366、403 个。值得注意的是,在胁迫时间点上,两个品种的差异代谢物数量都增加了,这表明了代谢产物对冷胁迫的积极适应。然而,CS 处理 7 天后,CT-C18 体内的差异代谢物含量高于 CS-C6,表明 CT-C18 具有较强的代谢紊乱。通过分析两个品种在不同时间点的差异代谢物类型,发现 CT-C18 品种对胁迫反应迅速,随着胁迫时间的延长,CT-C18 中上调的差异代谢物数量多于 CS-C6,而下调的差异代谢物数量与 CS-C6 相似。综上所述,CS- C6 未能累积到*多的差异代谢物,这可能与 CS 敏感性有关。相反,CT-C18 在整个应激条件下累积了较多的差异代谢物和较多的上调差异代谢物。这些数据表明,CS- C6 对冷处理敏感,而 CT-C18 在代谢组水平上对冷处理表现出显著的耐受性。
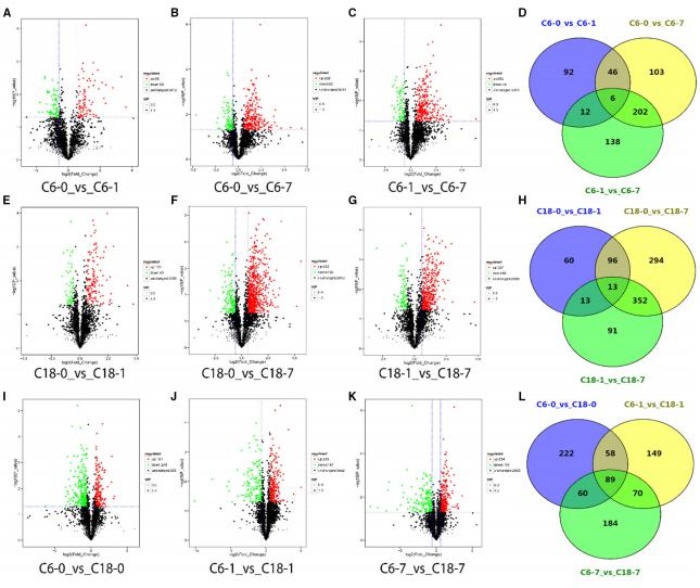
差异累积代谢物的火山图,以及对照与其时间点的比较
三、油菜品种的核心代谢产物对冷胁迫有反应
在 CS 条件下,在不同时间点对两个品种进行了共有和特有的差异代谢物的鉴定。然而,其中一些是已知的代谢物。因此,对命名的代谢物进行不同的比较。无论同一物种在不同的时间点或不同品种间的比较,都检测到相当大数量的差异代谢物。有趣的是,这些结果表明,与 CS-C6 相比,CT-C18 在同一时间点的比较中积累了更多的已知差异代谢物。KEGG 分析表明,这些代谢物主要属于以下几类:氨基酸代谢、碳水化合物代谢、其他次级代谢产物的生物合成、膜运输、脂类代谢、萜类和多酮类代谢、辅助因子和维生素的代谢、核苷酸代谢、能量代谢。
基于上述研究结果,本研究重点对两个品种在同一时间点检测到的已知差异代谢物进行了比较。有趣的是,CT-C18 品种的代谢产物比 CS- c6 品种的代谢产物表达上调,表明这些代谢产物在油菜籽对 CS 的耐受和适应中发挥了积极作用。根据 KEGG 注释,31 个差异代谢物涉及 19 条通路,包括生物合成通路、氨基酸代谢、碳水化合物代谢、核苷酸代谢、膜转运和能量代谢。
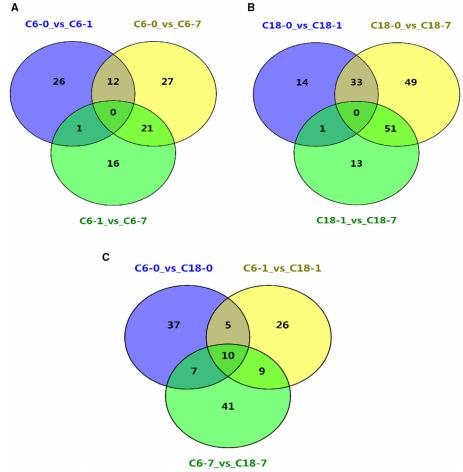
所有时间点共有的差异代谢物的维恩图
四、基于 CS 条件下 C6 和 C18 的转录组数据进行分析
在转录组分析中,检测到 C6-0 与 C18-0 比较中有 2845 个差异基因, C6-1 与 C18-1比较中有 3358 个差异基因, C6-7 与 C18-7 比较中有 2819 个差异基因。为了预测所有 DEG的功能,使用 MapMan 路径注释器对差异基因的代谢进行概述。油菜对 CS 的响应取决于其遗传背景。上调和下调的 DEG 表达趋势相似。主要表现在次级代谢,氨基酸,脂质、细胞壁、微量 CHO 和光反应途径。有趣的是,在应激 1 天和 7 天时,大多数途径都被发现富含上调基因。通过对油菜代谢途径的逐级综述,可以清晰地展现出油菜在 CS 处理后不同时间点的具体代谢途径。
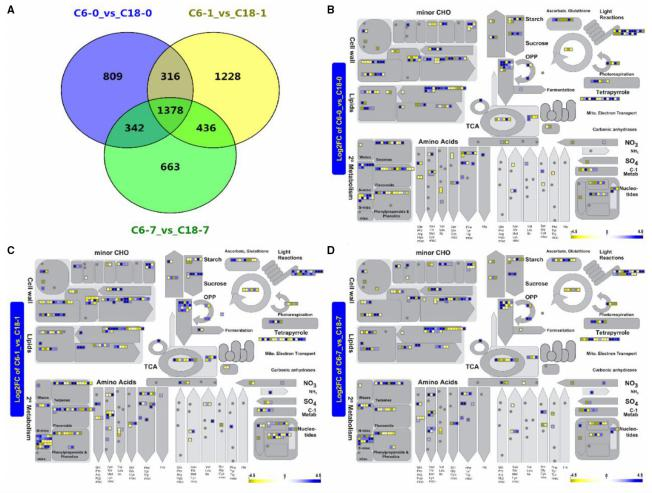
冷处理后不同时间点油菜作用的代谢途径
五、响应冷胁迫的差异代谢物和差异基因额调控网络和功能分析
为了了解 CS-C6 和 CT-C18 品种间的差异代谢调控网络,作者对 C6-0 和 C18-0、C6-1和 C18-1、C6-7 和 C18-7 三个组的核心差异代谢物与差异基因进行了相关性检验,并结合KEGG 数据库构建了调控网络。G1 调控网络分析表明,meta_908(海藻糖)和 meta_1297(N,N -二乙酰基壳生物糖)聚在一起,共享 19 个差异基因。然而,这些常见的差异基因在三种比较中表现出不同的表达模式。meta_56(咪唑油乙酸)、meta_190 (犬尿氨酸)、meta_1457(γ -生育三烯醇)和 meta_1678(角鲨烯)分别聚类。G2 调控网络分析表明,meta_217(4- 羟基苯基丙酮酸)与冷胁迫下第 0 天和第 7 天的差异基因相关。G3 调控网络分析表明,Meta_291(Alpha-D-葡萄糖)和 meta_481(脱氧腺苷)在胁迫第 1 天关联的差异基因数目高于第 0 天。G4 调控网络表明,meta_216(DL 香草甘露聚酸)和 meta_390(N-乙酰-l-苯丙氨酸)聚类,并与共有和特有的差异基因相关。总之,本研究结果表明,不同的差异代谢物和差异基因(参与相同的代谢途径)之间存在很强的正相关关系,从而促进了油菜对冷胁迫的耐受和适应性。
G1-G4 相关的差异表达基因进行 GO 富集分析和 KEGG 富集分析。发现富集的 GO-BP项主要是碳水化合物代谢过程、细胞氨基酸代谢过程、有机酸代谢过程、羧酸代谢过程,氧合酸代谢过程。氨基酸和核苷酸、淀粉和蔗糖的代谢的代谢主要富集于不同时间点 0, 1 和 7天。KEGG 富集分析中,G1 的结果表明,0、1 和 7 天时,主要富集在氨基酸和核苷酸的糖代谢。淀粉和蔗糖的代谢也有类似的趋势。G2 的结果显示,在冷处理的 1 天和 7 天两个时间点上,氨基酸的生物合成是*显著和富集的途径。值得注意的是,2-氧羧酸代谢在冷处理的第 7 天特异性富集。G3 的结果表明,在冷处理的 0 和 1 天,碳代谢富集,这与相应的GO-BP 分析一致。同样,在 CS 第 1 天,苯丙氨酸代谢、酪氨酸代谢、异喹啉生物碱生物合成、曲烷、哌啶和吡啶生物碱生物合成均富集。GO-BP 和 KEGG 富集分析表明,主要氨基酸和碳水化代谢在油菜 CS 耐受性和适应性中起关键作用。
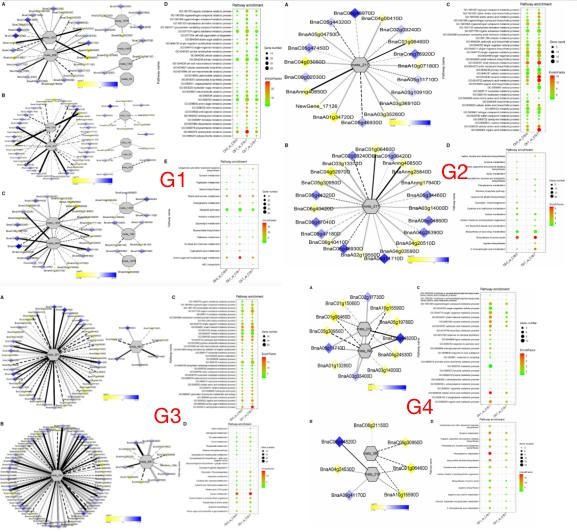
注:低温胁迫相关基因与 dam 的相关网络分析。G1:“C6-0_vs_C18-0”、“C6-1_vs_C18-1”和“C6-7_vs_C18-7”共有的 10 个已知差异代谢物;G2:“C6-0_vs_C18-0”和“C6-7_vs_C18-7”共有的 7 个已知差异代谢物;G3:“C6-0_vs_C18-0”和“C6-1_vs_C18-1”共有的 5 个已知差异代谢物;G3:“C6-1_vs_C18-1”和“C6-7_vs_C18-7”共有的 9 个已知差异代谢物。
六、淀粉和蔗糖代谢对 CS 的反应
通过 GO 和 KEGG 富集分析表明,在冷胁迫处理下,油菜的许多差异代谢物和差异基因与淀粉和蔗糖代谢相关。海藻糖和纤维二糖在胁迫过程中积累,表明 CS-C6 应保持高于 CT-C18 的海藻糖和纤维二糖水平。同时,糖原、D-葡萄糖-6P 和 D-果糖-6P 在处理的不同时间点均被下调,说明相比 CS-C6,CT-C18 应保持较高的水平。热图中总结了参与淀粉和蔗糖代谢途径的差异基因的表达模式,这些差异基因与上述 5 个差异代谢物中的 1-3 个相关,并在两个品种中均显示出不同的表达模式。值得注意的是,与葡萄糖分支直接相关的三个差异基因在C6中的表达水平较低,随后导致d-葡萄糖-6p在CS-C6中的积累水平较低,其他的糖类变化并不明显。以上结果表明,高含量的糖相关化合物对油菜的 CS 耐性有显著的促进作用。在冷处理条件下,大多数负责碳水化合物生物合成的基因都表达上调,并且在暴露后 1 或 7 天,CT-C18 的变化大于 CS-C6。这表明CT-C18 在积累一些碳水化合物方面表现更好,因此,与 CS-C6 品种相比,表现出更高的 CS 耐受性水平。相反,这些分子水平较低可能导致冷敏感性。
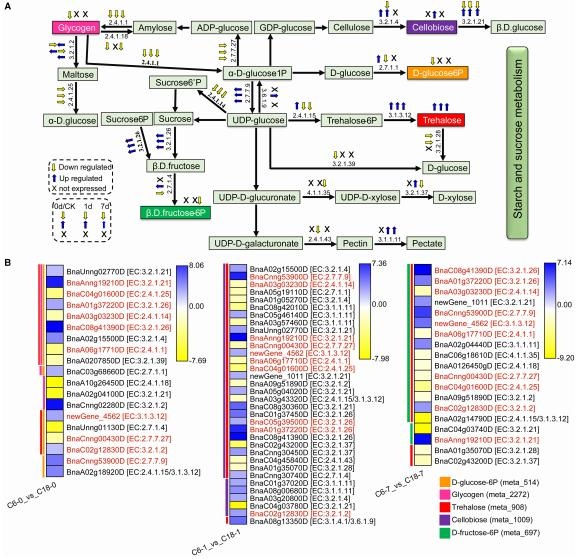
C6 和 C18 品种响应 CS 的淀粉和蔗糖代谢途径
七、苯丙氨酸代谢对 CS 的反应
苯丙氨酸代谢在代谢组和转录组数据中都显著富集,值得注意的是,在第 0 天和第1 天,只有 2-苯乙酰胺和 n -乙酰- l-苯丙氨酸积累,而在第 7 天,2-羟基苯乙酸、n -乙酰- l-苯丙氨酸、3-苯基丙酸和水杨酸的含量上调,琥珀酸的含量下调。参与苯丙氨酸代谢途径的差异基因的表达模式见热图(图 10B)。特别是,通过 3 次比较检测到 5 个共有差异基因。其中三个分别负责苯丙氨酸或苯丙酮酸和反式 4-羟基肉桂酸的生物合成的基因表达下调。三次比较中,少数基因表达上调(图 10A),表明 CT-C18 中差异基因的表达水平较高。而苯丙氨酸代谢的其他最终产物和中间产物,除 3 个差异代谢物外,在两个品种之间没有差异。这些结果表明,油菜籽通过苯丙氨酸代谢来调节氨基酸的积累。然而,这些水坝的较高含量,主要是在 CT-C18 中,可能与油菜籽的 CS 耐受有关。
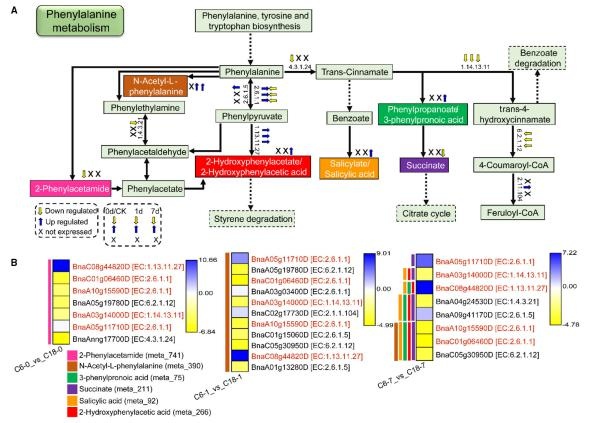
在 C6 和 C18 品种中响应 CS 的苯丙氨酸代谢途径
结论
通过结合转录组和代谢组数据分析,作者发现大量的差异代谢物与几个差异表达基因密切相关。通过功能富集分析表明,大部分的差异基因和差异代谢物主要富集在不同的碳水化合物和氨基酸代谢中。其中淀粉、蔗糖代谢和苯丙氨酸代谢显著丰富,对油菜冷胁迫适应起着至关重要的作用。从低温适应控制的两条通路中筛选出 6 个候选基因。在进一步验证的基础上,对拟南芥的同源 T-DNA 插入突变体 4cl3, cel5, fruct4, ugp1,axs1, and bam2/9进行了鉴定,发现 6 个突变体的抗冻性存在显著差异。本研究结果为了解油菜低温胁迫响应的分子基础和耐受机制提供了新的前景,并为提高油菜低温胁迫适应性提供了一组候选基因。




 京公网安备 11011302003368号
京公网安备 11011302003368号