
发表期刊:Neuron
影响因子:17.173
原文链接:https://doi.org/10.1016/j.neuron.2020.12.010
发表日期:2021-01-06
摘要
单细胞测序技术,包括转录组和表观基因组分析,正在改变我们对神经回路的细胞构建模块的理解。单细胞测序方法可以通过直接测序数千到数百万单个细胞中的多个分子信号,全面表征脑细胞类型的多样性。这些测序结果可以揭示特征细胞的基因调控机制,并提供脑细胞种群之间发育和进化关系。单细胞测序数据可以帮助设计用于脑电路组件的目标功能研究的工具,将分子特征与解剖学、连通性、形态学和生理学联系起来。在此,作者讨论了单细胞转录组和表观基因在神经科学中的关键应用。
前言
脑细胞是复杂的生物机器,其功能是由保守的基因表达程序在分子水平上定义的。要了解神经细胞类型的分子特征,需要测序数千个基因,以发现细胞之间的细微差别。与此同时,需要大量的细胞样本来捕获罕见类型并准确评估他们之间的差异。对大量基因的细粒度分析和对大量细胞的覆盖率的广度分析常常被视为相互排斥的目标:通常情况下,实验往往是在有限的样本中检测尽可能多的分子特征,或在大量样本中瞄准少量基因。在本文中,作者对高通量单细胞测序分析是如何实现高分辨率、宽范围的脑细胞类型分析进行总结和阐述。
DNA和RNA测序量化信息承载分子编码并且制定每个细胞的生物学特性。RNA分子的丰度属于转录组研究,而DNA和组蛋白的化学修饰以及细胞核内DNA的物理构象则属于表观基因组范畴(图1A)。
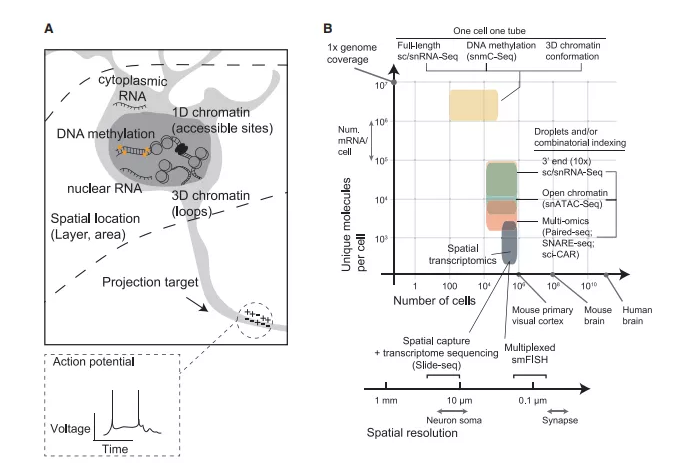
图1:神经科学中的单细胞测序方法
成千上万的基因和成千上万的基因调控序列之间的相互作用,在每个细胞中产生了数百种的细胞表型。神经科学家致力于利用细粒度、大范围的单细胞测序数据来促进对脑细胞类型的转录组和表观基因组特征的研究。
单细胞测序样品的细胞信息携带分子
目前,对脑组织中RNA和DNA的分析在很大程度上还局限于对组织平均成分的批量分析。批量分析破坏了转录本或DNA修饰与单个细胞的联系,可能无法检测到罕见的细胞类型。一种细胞类型的基因表达特征可能被其他细胞类型的互补模式所掩盖。尽管从特定细胞群纯化整个细胞或细胞核的技术可以部分克服这些限制,然而,这需要高度选择性的RNA或蛋白质标记或遗传工具(如小鼠CRE系),并且只能应用于先前已确定的细胞类型。
单细胞测序技术对来自单个细胞的RNA或DNA进行测序,不需要选择性的细胞纯化。这些技术可以概括为三个特征:范围(细胞数量)、粒度(基因或表观遗传特征的数量)和空间分辨率(图1B)。此外,单细胞技术可以检测不同类型的RNA转录本或表观遗传修饰。虽然单细胞测序不需要细胞纯化,但它依赖于高质量的原组织或冷冻组织。
哺乳动物细胞的转录组由10^5-10^6个单独的信使RNA (mRNA)分子组成。这些信息代表每个细胞约4000 ~ 12000个不同的基因,包括许多编码基因的多个变异的不同的mRNA亚型。尽管转录后调控是复杂的,但细胞中mRNA转录本的数量与编码蛋白的丰度相关,因此是细胞身份和功能的标志。
单细胞表观基因组分析方法还可以用于检测DNA甲基化、染色质可及性和染色质构象等。这些DNA的化学和物理修饰是在发育过程中建立起来的,并在整个生命周期中调节基因的表达。而转录组信息表现了基因表达的差异,部分反映了细胞的状态和组织收集前后神经活动的影响,表观遗传标记(包括长寿命染色质成分的短暂和稳定的修饰)。表观基因组数据还可以解析顺式调控元件的功能,如增强子可以建立和维持细胞类型特异性的基因表达。
综上所述,单细胞转录组和表观基因组测序共同构成了解析大脑细胞成分的强大工具,使研究健康和疾病大脑中特定细胞类型的功能成为可能。
单细胞转录组学
单细胞mRNA测序技术(scRNA-seq)使得脑细胞类型分子研究进入了的新时代。与常规的RNA测序(RNA-seq)一样,scRNA-seq以mRNA分子为模板进行逆转录合成互补DNA (cDNA)。二代测序(NGS)文库是通过cDNA扩增、片段化、附加扩增和NGS定量等步骤生成的。
多孔板法和液滴法
单个细胞或细胞核可以通过基于多孔板的细胞分选或液滴分选进行物理分离。基于多孔板的分选方法使用荧光激活细胞分选(FACS) 或显微镜引导的毛细管将单个细胞或一组细胞分布到不同孔中。基于液滴的方法将细胞分离到脂质悬浮的单个水室中。细胞在液滴中被裂解,mRNA分子被逆转录,并用能识别细胞的寡核苷酸条形码标记。基于液滴的scRNA-seq是高效的,因为它的反应体积小,通过微流体装置的连续流动可以快速处理数千个细胞。
通过物理方式分离细胞的另一种方法是使用细胞特异性条形码组合的cDNA多路测序。一组细胞被分配到孔中(池化),用特定于孔的条形码标记,然后合并,重新分配到不同的孔中,并用另一种条形码标记。经过多次池化、分离和标记,细胞获得随机的条形码组合。可以调整每个孔的单元数,以确保大多数单元具有独特的条形码组合,由于条形码碰撞可能会发生一些重叠。然后将来自所有细胞的cDNA汇集在一起并进行测序,然后通过计算将数据解析出来。基于不同条形码组合和液滴的方法具有更高的通量和更低的单位细胞成本,而基于平板的方法提供更敏感的基因检测和更可定制。
重要的是,这些策略在量化mRNA丰度方面存在差异(图2A)。基于液滴的scRNA-seq方法计数方法对mRNA分子的5’ 端或3’ 端进行计数,结合独特的分子标识(UMIs),以避免重复计数同一分子的PCR产物。这些数据并不能区分共享相同启动子或转录终止位点的mRNA异构体。相比之下,基于多孔板的分选策略可以对全长转录本的片段进行测序,包括覆盖所有表达外显子和剪接连接的内部片段。这些方法表明mRNA异构体的使用在不同的脑细胞类型中是不同的。通过结合UMIs的全转录序列测序,可以在单细胞中重建数千个独特的mRNA分子。
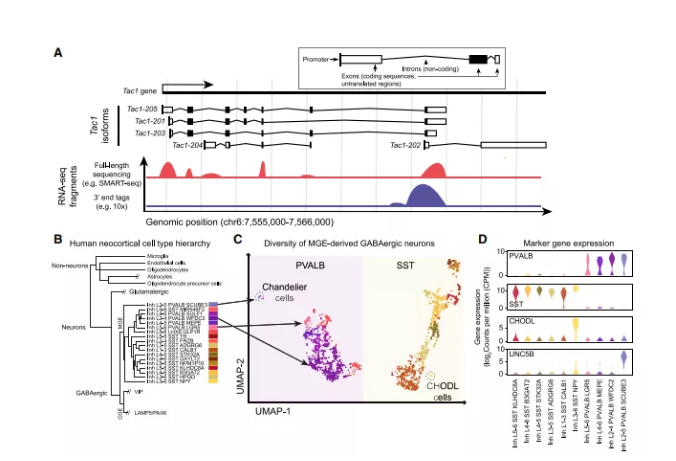
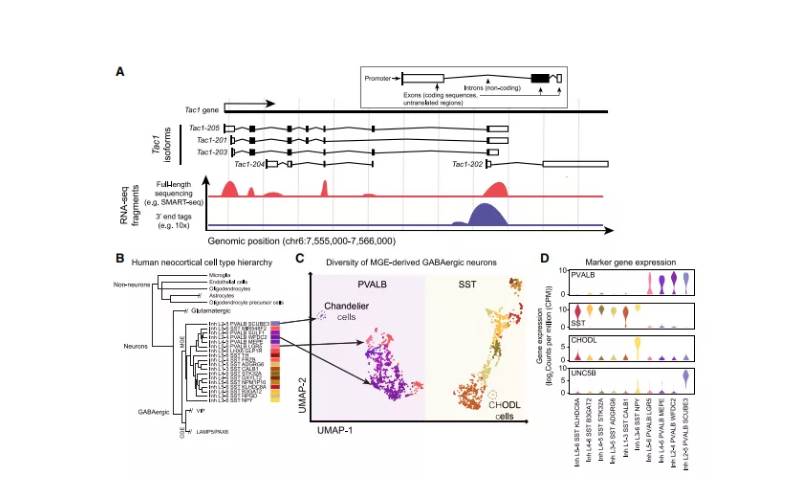
图2:单细胞转录组学在神经科学中的应用
单细胞和单细胞核
脑组织中树突、轴突和胶质过程的复杂网络对scRNA-seq提出了多重挑战。远端细胞中包含大量的能够指导合成树突状蛋白的mRNA。然而,这些转录本在解剖和细胞分离后大部分丢失。因为所有转录本都起源于细胞核,scRNA-seq仍然可以在细胞核或体细胞中捕获新合成的树突和轴突mRNA,然后将其运输到远端细胞中。此外,scRNA-seq不能应用于冷冻的脑组织,因为在冷冻过程中细胞膜会破裂。
另一种方法是单细胞核测序。核转录本包含细胞中的20%-50% RNA,包括未成熟RNA以及未剪接RNA分子(图2A)。尽管如此,与scRNA-seq相比,snRNA-seq提供了更好的脑细胞类型标记和分辨率。
与scRNA-seq相比,snRNA-seq数据可能较少受到某些技术限制的影响。由于细胞核的大小和形态相对一致,snRNA-seq比scRNAseq更有可能捕获所有类型的细胞。此外,在组织剥离和细胞分离过程中,snRNA-seq不太容易受到虚假的基因表达激活的影响。
单细胞转录组学在神经科学中的应用
细胞类型定义和表征
单细胞技术改变了我们对脑细胞种类多样性的认识。几十种形态和功能上截然不同的脑细胞类型已经被确认,但是传统的显微镜和生理学方法对哺乳动物大脑回路中数百万个细胞的细胞类型进行全面表征的范围有限。最近的scRNA-seq研究在小鼠大脑区域取样可以达到20,000到 750,000个细胞。最大的单个scRNA-seq/snRNA-seq数据仅代表了成年小鼠大脑中近1亿个神经元和胶质细胞的1%(图1B)。
人类大脑的神经元细胞比小鼠大脑多1000倍,全面取样更令人生畏。来自人类大脑多个区域的单核转录组已经建立了细胞类型分类(图2B-2D) 。将人类与小鼠和非人类灵长类动物进行比较,揭示了保守而独特的细胞类型特征,包括细胞群体比例的物种特异性差异,以及同源细胞类别和类型内的基因表达的差异。scRNA-seq/snRNA-seq还提供了对胶质细胞多样性的了解,包括少突胶质细胞谱系和小胶质细胞的功能状态。
scRNA-seq也被应用于非哺乳动物的大脑,阐明了关于发育、细胞调控和衰老的基本问题。例如,衰老极大地减少了果蝇脑细胞中的RNA数量。在斑马鱼的大脑中,不同的神经干细胞群体在阿尔茨海默病(AD)模型中受到淀粉样蛋白毒性的影响不同。
发展和可塑性
scRNA-seq揭示了小鼠和人类大脑甚至在整个生物体发育中的神经元和胶质细胞类型的发育。scRNA-seq/snRNA-seq是特别适合研究大脑发育的方式,因为关键的干细胞和神经祖细胞群可能是短暂的,在没有事先了解特定分子标记的情况下很难分离。scRNA-seq在多个时间点可以重建发育轨迹,揭示转录因子的动态和下游效应体调节特定的神经群体。新生神经元和成人少突胶质细胞的分化轨迹也可以重建。一个scRNA-seq/snRNA-seq的前沿领域是与人工标记的谱系追踪结合或与一个谱系内细胞自然的体细胞突变相结合。
除了发育动力学外,神经活动和可塑性导致的基因表达变化可以通过snRNA-seq/scRNA-seq评估,如视觉皮质神经元在光照后的表现。此外,分析活性调节基因可以识别活性细胞。考虑到活性和钙信号依赖的基因调控在突触可塑性中的关键作用,体系的背景信号对于单细胞计数分辨细胞类型特异性的转录组动力学至关重要。
细胞类型在疾病中的作用
通过应用于患者死后脑组织的snRNAseq,可以研究疾病的转录组特征。通过对48例AD患者的80000个单核细胞进行snRNA-seq检测,在单个细胞类型中检测到的差异表达(DE)基因比在组织中检测到的多。特别是,神经胶质细胞的基因失调在snRNA-seq中检测到,但在大量的RNA-seq中没有检测到,因为神经胶质对组织中总RNA的贡献相对较低。snRNA-seq已经应用于AD、重度抑郁症、自闭症、雷特综合征和多发性硬化症等疾病患者的脑组织。
值得注意的是,疾病研究集中在相对少量的转录组或表观基因组簇对应于细胞类别,而不是单个类型。解剖细粒细胞类型对疾病的贡献是重要的,但具有挑战性,因为在计算分配单个细胞集群的偏差可能会显著影响研究结果。
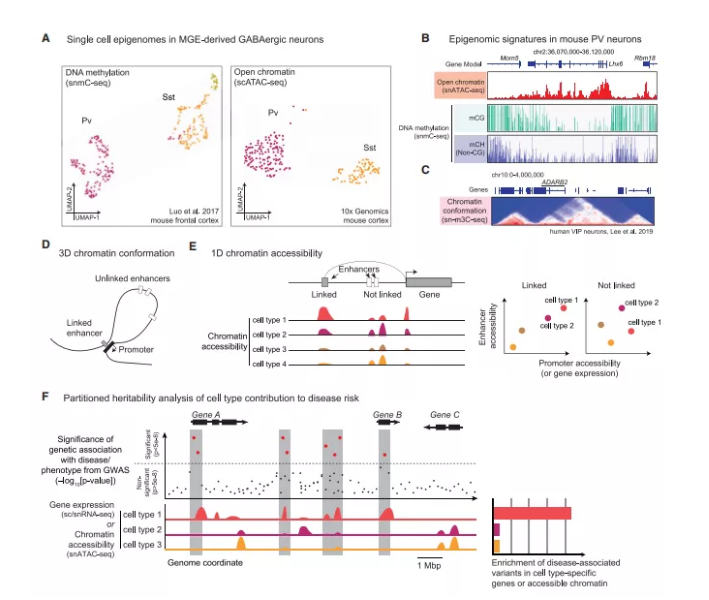
图3:单细胞表观基因组学
单细胞表观基因组学
基因的稳定表达部分是通过DNA的表观遗传修饰来维持的,如基因组胞嘧啶甲基化和组蛋白的甲基化修饰。这些标记通过引导转录因子蛋白与特定基因组区域的结合来影响基因表达,从而增强或降低附近或远端基因的转录。尽管单个mRNA分子半衰期为几分钟到几小时,但有丝分裂后的神经元中DNA和某些组蛋白的共价修饰可以持续数月到数年。表观基因组的动态调节跨越脑细胞类型和整个生命周期,建立和维持细胞身份,并且可能影响染色质可塑性和行为。单细胞表观基因组可以补充转录组,提供细胞类型特异性基因表达调控的方式。
与snRNA-seq一样,表观基因组分析可以应用于从冷冻和储存的死后组织中获得游离的细胞核。这些方法可以评估包括超过95%基因组的非蛋白质编码区域。非编码区包含数百万个影响基因表达的候选顺势作用元件(cCREs),包括启动子和增强子。cCREs的定义是染色质开放区,DNA甲基化水平低,并且可以通过细胞核内DNA的三级结构与基因的启动子物理相互作用。识别细胞类型特异性的增强子可以指导针对功能研究细胞类型的病毒载体和转基因株系的开发。未来,我们将在单细胞水平上研究表观基因组的动态调控,包括发育轨迹和学习和记忆过程中神经元活动依赖的基因组修饰。
DNA甲基化
在哺乳动物中,基因组胞嘧啶可以被一个甲基基团共价修饰。甲基胞嘧啶(mC)经常在CG双核苷中被发现,它经常与转录抑制相关。与其他细胞相比,脑细胞中的DNA甲基化有两个独特的特点。首先,神经元具有高水平的羟甲基胞嘧啶(hmC),这是mC的衍生物,可能具有不同的功能。其次,在发育过程中,神经元在非CG位点积累大量mC,(主要是CA和CT二核苷酸)。在整个基因组中,近10亿个CG和非CG位点的DNA 甲基化修饰(mC)和羟甲基化修饰(hmC)是神经元和胶质细胞中细胞类型特异性基因调控的特征。
DNA甲基化可以使用基于多孔板的单个核分选,然后进行DNA的亚硫酸氢盐转化和测序,每个细胞可以捕获高达30%的基因组。亚硫酸氢钠处理可以将胞嘧啶转化为尿嘧啶,而甲基化的胞嘧啶无法被转化,因此在DNA亚硫酸氢盐转化后进行测序可以揭示甲基化状态胞嘧啶位置。该技术从45个小鼠大脑区域获得了超过110,000个单细胞DNA甲基化信息,在大脑皮层、纹状体和嗅觉区域识别了161个准确的神经元细胞类型。每个神经元群体在基因体上都有独特的DNA甲基化特征,并且这些特征与基因表达密切相关(图3B)。基于液滴的亚硫酸氢盐化学可以潜在地提高单细胞DNA甲基组分析的效率,但与转录组测序相比,全基因组测序的高成本仍将是限制该技术在大量细胞中应用的一个因素。未来,单细胞的羟甲基化分析将阐明神经元细胞类型中mC和hmC的相对数量。
一维染色质可及性图谱
正如基因是转录组分析的基本单位一样,cCREs是表观基因组分析的基础。cCREs可以通过与转录因子蛋白结合导致核小体移位,形成可及性染色质区域。snATAC-seq可以利用Tn5转座酶捕获并绘制这些区域的图谱。
与scRNA-seq/snRNA-seq一样,snATAC-seq也可以使用液滴及index组合进行分析。snATAC-seq可应用于冷冻脑组织,每次实验可以捕获大于10^4个细胞。值得注意的是,在所有可以在特定细胞类型中检测到的远端cCREs中,snATAC-seq捕获大约每个细胞的2%-3%。这种较低的覆盖率可以反映该试验的敏感性,和在同一类型的单个细胞间的异质性可及性。
尽管覆盖率较低,snATAC-seq数据可靠地准确区分了神经元和胶质细胞类型。例如,我们使用snATAC确定了160种细胞类型。在所有类型的细胞中,总共发现了50万个可达区域(cCREs)。另外,在用Tn5转座酶批量标记开放的染色质后,液滴可用于物理分离细胞核并进行标记,每个细胞产生多达34,000个独特的测序片段。虽然scRNA-seq/snRNA-seq数据需要使用先前注释过的基因和转录本进行量化的,但可获得的染色质区域在整个基因组中分布,通常无法提前推断。许多类型的特征已被用于量化snATAC数据,包括基因启动子、假定的增强子以及共同DNA序列基序的区域组合等。每一种方法对不同的生物信号都很敏感,并受到片段采样所引入的背景信号的不同影响。计算分析方法的系统基准和比较对于实现最大可能的分辨率很重要。
染色质的三维构象
CREs和基因启动子通过染色质环和三维结构域相互作用(图3D)。染色质三维构象的单细胞分析与在大量组织中使用的高通量基因组捕获技术(Hi-C)密切相关,通过交叉连接、碎片化和在位置上接近的DNA片段重组,识别相互作用的区域。对得到的文库进行测序可以检测到嵌合DNA片段,这些片段包含一维基因组中相隔很长一段距离,但它们在三维空间中紧密相连的序列。这些关联的信息可以用接触矩阵表示(图3C)。
染色质三维构象分析的一个挑战是染色体位置之间有很大的空间可以发生可能的成对接触。即使在单细胞Hi-C实验中可以捕获基因组的很大一部分,产生的成对接触矩阵也只包含基因组位置三维之间接触的一小部分,因为每个DNA片段只能反应一次连接事件。单细胞Hi-C数据的聚类分析可以识别具有相似染色质组织的细胞组,但其本身并不能可靠地区分皮质神经元类型。一种替代策略是使用薄的(- 0.2mm)组织切片,然后用激光捕获单个核和同一切片中的测序片段。
单细胞表观基因组学的应用
解剖神经元基因调控网络
单细胞表观基因组可以用来描述有关建立和维持脑细胞类型识别的转录因子和cCREs之间的调控网络。与DNA结合的蛋白通常识别6-10个碱基对的特定序列或基序(图3A,Luo 等,2017)。通过检测cCREs中富集的序列,并与已知的转录因子结合基序进行比较,可以识别调节特定脑细胞类型的转录因子。该方法应用于整合小鼠运动皮层的多模态(转录组和表观基因组)数据,揭示了转录因子Rfx3在调节兴奋神经元中的作用。通过检测结合位点的染色质可及性特征,可以对转录因子结合的确切位置进行高精度分析。然而,到目前为止,这种分析仅限于大量样本的染色质可塑性分析。
遗传因素与疾病风险的联系
尽管全基因组关联研究(GWASs)已经确定了一些神经和精神疾病的风险位点,但每种疾病中特定脑细胞类型的脆弱性在很大程度上仍是未知的。通过细胞类型特异性转录组或表观基因组的遗传分析可以识别活跃基因在GWAS风险位点附近富集的细胞类型,表明其在疾病中可能具有直接或间接的作用(图3F)。
局限和展望
单细胞测序正在改变生物学的许多领域,其影响可能在神经科学方面尤其深刻。脑细胞类型的多样性,以及在整个神经元生命周期中细胞身份和依赖经验的可塑性的复杂调节,使得单细胞分析对于理解大脑回路至关重要。
像其他任何新兴技术一样,单细胞测序也有重要的局限性。在单孔或液滴中捕获两个或多个细胞,或用相同的条形码标记,可能会使单细胞数据不准确,并产生虚假的混合细胞类型。RNA-seq文库的污染会导致假阳性结果。更大的局限性在于假阴性或丢失,因为采样策略通常只恢复总RNA或DNA的一小部分。单细胞技术也会受到组织剥离和细胞分裂引起的非生理转录的影响。
单细胞技术的一个更基本的限制是需要破坏细胞以提取其分子特征以进行测序。虽然多组学和多模态技术可以增加每个细胞获得的信息,但这些方法本质上是细胞生命周期的快速捕捉,并没有记录对大脑功能至关重要的神经调节的动态变化。通过分析成熟和不成熟的RNA转录本,可以推断出一些动态信息,但在单细胞中真正测量转录组动力学将在大脑相关的研究中有重要价值。
单细胞数据集的复杂性既是机遇也是挑战。用于聚类、可视化和分析单细胞数据的复杂计算方法为深入了解细胞的功能提供了丰富的见解。然而,对于相同的数据,不同的计算方法或参数选择可能导致不同的结果。聚类程序因其不一致性和缺乏稳定性而导致在描述单细胞数据的细胞类型方面的主观性。单细胞研究必须通过公开透明地调查和生物学结论报告如何依赖于分析选择,并公布代码和数据来重现所有分析解决这一问题。
最大的挑战是将来自单细胞测序的信息与神经元细胞类型识别的功能和生理指标统一起来。尽管在转录组学、表观基因组学、解剖学和形态学标准定义的细胞类型之间存在显著的对应关系,但在数百种精细脑细胞类型水平上协调这些数据仍处于早期阶段。
随着单细胞技术的创新克服了这些挑战,它们正在为神经科学研究开辟新的前沿领域。单细胞测序提供了对多个物种和不同发育阶段大脑细胞成分的全面、全脑分析。将单细胞测序与电生理学、形态学和连通性联系起来的技术将深刻地改变我们对神经元多样性维度的理解。跨物种和发育阶段的脑细胞类型的比较将改变我们对大脑发生和系统发育的理解,并可能影响对神经群体间进化关系的详细机制理解。进化保守性和分化提供了对基因调控程序的深入了解,这些基因调控程序对因为它们认知功能的影响而被选择。通过结合单细胞测序和活性依赖标记技术,神经科学家将能够将转录组和表观基因组与特定细胞类型在行为中的作用联系起来。除了技术进步,我们期望新的概念和理论的出现,从而将从单细胞测序的巨大信息转化为关于脑细胞个体发生和功能的有组织和系统的知识。
推荐阅读
◆ 单细胞多组学揭示anti-PD-L1免疫治疗联合化疗在三阴性乳腺癌中应答关键的免疫细胞亚群
◆单细胞多组学|Nature带你玩转单细胞表观基因组学探讨人类皮质发育的机制
◆空间单细胞转录组研究成果 | 人类乳腺癌的时空图谱-细胞表型和肿瘤亚型
◆【单细胞+空间转录组】单细胞和空间转录组强强联手共同研究人类肌腱疾病中免疫稳态失调




 京公网安备 11011302003368号
京公网安备 11011302003368号