

中文名:通过lncRNA-mRNA共表达网络分析长非编码RNA调节蝗虫的型变
英文名:Long Non-coding RNA Derived from lncRNA– mRNA Co-expression Networks Modulates the Locust Phase Change
期刊:Genomics Proteomics Bioinformatics
IF:7.051(1区)
通讯作者单位:中科院、河北大学
研究背景
长链非编码RNA(lncRNA)调节基因表达、动物行为等各种生物学过程。尽管蛋白质编码基因、microRNA和神经肽在亚洲飞蝗的表型可塑性调节中起着重要作用,但有关lncRNA在此过程中功能研究较少。本文应用高通量RNA-seq来比较蝗虫型变时程中lncRNA和mRNA的表达模式。结果显示lncRNA在型变的早期阶段反应更快。功能注释表明,早期改变的lncRNA在分散和群居阶段采用了不同的途径来应对种群密度的变化。筛选了分散和群居阶段的网络中两个重叠的中枢lncRNA基因座进行功能验证。本文进一步证明LNC1010057为潜在的蝗虫型变因子。这项工作为深入了解蝗虫型变的分子机制并扩大lncRNA在动物行为中的作用范围提供了重要的数据。
材料方法
实验材料:散居化处理为将群居型蝗虫单独饲养0h、4h、8h和16h后取出手机蝗虫大脑进行测序;
群居化处理为将10只散居型蝗虫与20只群居型蝗虫饲养于一个小笼子(10厘米×10厘米×10厘米)中,于0h、4h、8h和16h后取出,收集蝗虫大脑进行测序。在同一时间点采集样品的三个生物学重复,提取RNA,建立cDNA文库并进行RNA-seq。对lncRNA和mRNA进行差异表达分析及qRT-PCR验证,对不同时间点的样本进行STEM(ShortTime-seriesExpressionMiner)分析,并构建lncRNA–mRNA共表达网络的构建。对目标中心节点lncRNA进行RNAi,采集其行为录像数据进行行为数据分析。
实验结果
1、蝗虫lncRNA的外显子比mRNA少但更长
研究发现大约78%的lncRNA包含2个外显子,而mRNA中包含的外显子数量为1至120(图1C)。因此,lncRNA的外显子明显长于mRNA的外显子(平均长度1142bpvs.263bp,图1D,左)。同时,lncRNA的内含子明显短于mRNA的内含子(平均长度:10086bpvs.12442bp,图1D,右)。表达水平分析表明,lncRNA的总体表达水平明显低于mRNA的表达水平(平均值为0.6vs.1.9;图1E)。但是,表达特异性分析表明,lncRNA的表达受时间限制的程度要高于mRNA(平均值为0.672对0.436图1F)。基于相对于mRNA的lncRNA基因组位置,蝗虫lncRNA被分类为基因间、重叠区、有义内含子、反义内含子、有义外显子和反义外显子。蝗虫77%以上的lncRNA是长基因间的ncRNA(lincRNA,图1G)。这些结果表明,蝗虫lncRNA与mRNA在结构和表达上有很大不同。蝗虫lncRNAs更长,具有更少但更长的外显子和更短的内含子。此外,lncRNA的表达模式显示出比mRNA更高的时间特异性。
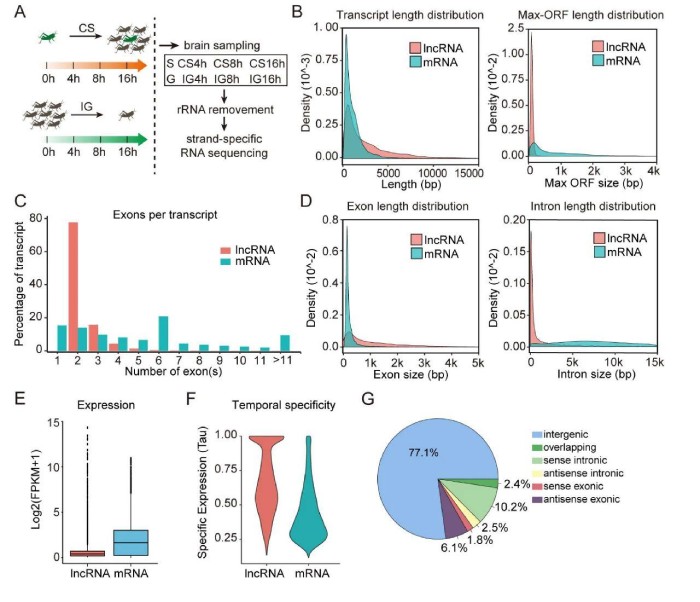
图1lncRNA和mRNA之间的不同结构和表达
2、群居型蝗虫中特异性表达的lncRNA比在散居型蝗虫中表达的更多
在散居型和群居型蝗虫大脑中共表达9722个lncRNA(73.9%),而散居型蝗虫中特异性表达962个lncRNA(7.3%),群居型蝗虫中特定表达2469个lncRNA(18.8%)(图2A,顶部)。分别在散居型和群居型蝗虫中特异性表达了479和1090个mRNA(3.1%和7.1%)(图2A,底部)。这些结果表明,与mRNA相比,在两个蝗虫相型特异性表达的lncRNA的百分比更高,而在群居蝗虫中比散居型蝗虫中表达的lncRNAs更多。与散居型蝗虫相比,群居型蝗虫中335个lncRNA和779个mRNA的表达水平下调,而313个lncRNA和261个mRNA的表达上调[倍数变化(FC)>2和P<0.05;图2B]。lincRNA在下调和上调的lncRNA中所占的比例高(分别为81.2%和87.8%),其次分别是反义外显子和有义内含子lncRNA(图2B)。在表达中按FC排名的前10个lncRNA基因显示在图2C中。这表明散居型和群居型蝗虫大脑中表达的lncRNA的数量及其表达水平明显不同。
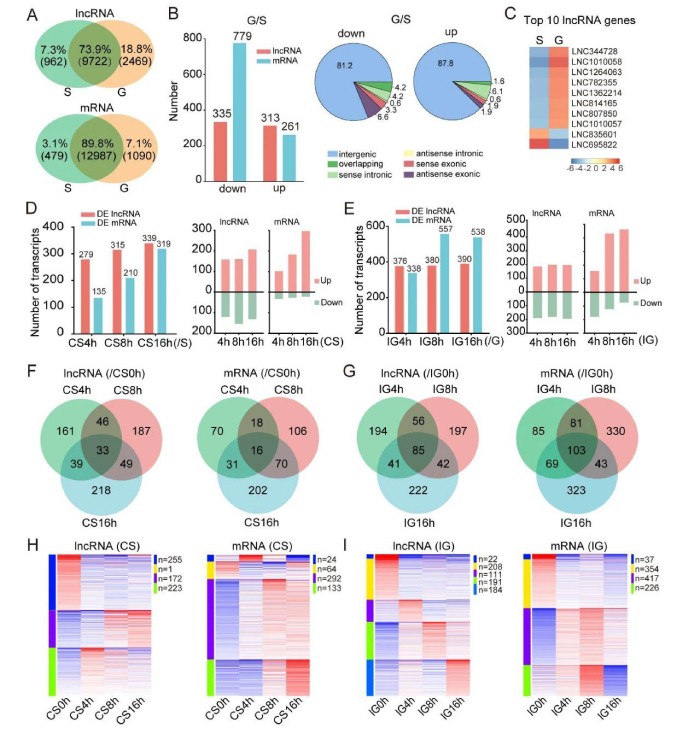
图2lncRNA在蝗虫型变中显示出不同的表达变化模式
3、lncRNA对种群密度变化的快速应答
为了进一步分析lncRNA和mRNA的表达模式,使用了短时间序列表达挖掘器(ShortTime-seriesExpressionMiner,STEM)对基于表达的转录本进行聚类。在CS期间,将214个lncRNA和242个mRNA分别分为8个和9个重要的表达谱(图3D)。其中lncRNA表达谱15以及mRNA表达谱15、13和16直到群居化处理后8或16小时才改变。这些配置文件称为后期更改的配置文件(模式d)。与后期更改的配置文件相反,早期更改的配置文件以4小时时间点开始的转录表达变化为特征。根据4小时后的表达变化,将早期变化的轮廓细分为模式a(早期变化)、模式b(早期中变化)和模式c(可持续变化)。在IG过程中,269个lncRNA和639个mRNA分别聚集成6个和7个显着表达谱。所有lncRNA配置文件均为早期变化(图3E)。在mRNA表达谱中,有6个是早期改变的,而谱12是晚期改变的。在CS和IG期间,lncRNA的聚类分布图的数量与mRNA的分布无明显差异。但是,通过计算谱图中的转录本数量,发现早期改变的lncRNA的百分比比CS中的mRNA的百分比高61.1%(88.3%vs.54.8%,图3F)。同样,IG中早期改变的lncRNA的百分比高于IG中的mRNA(100%vs.81.3%,图3F)。因此,lncRNA比mRNA对散居化和群居化处理的反应更快。在CS中,早期改变的lncRNA的表达变化速率快于285个mRNA的表达变化速率(0.55vs.0.24)和IG(0.77vs.0.39)(图3G)。因此,早期改变的lncRNA的比例和表达变化率高于早期改变的mRNA。因此,蝗虫lncRNA比mRNA对种群密度的变化更敏感。
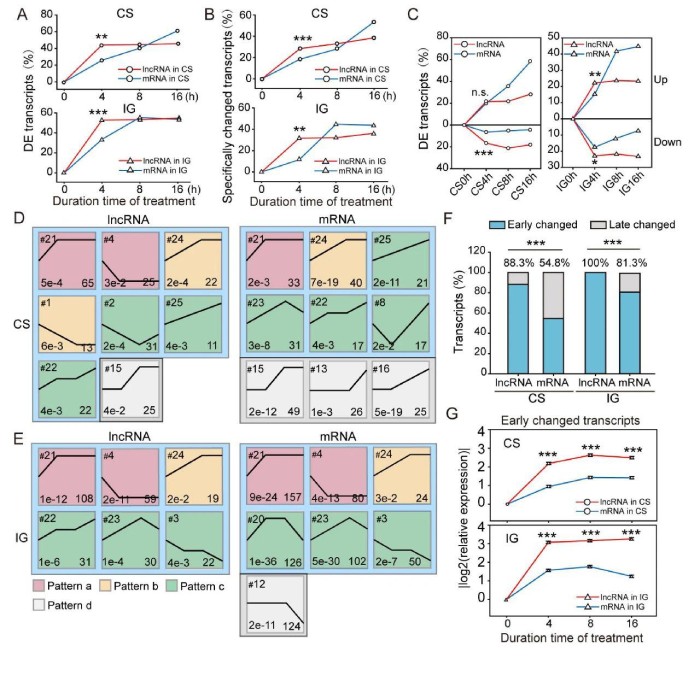
图3lncRNA对散居化和群居化处理的快速反应
4、lncRNA参与CS和IG早期变化的不同途径
在CS中,自噬调节、剪接体和肌醇磷酸代谢途径在上位和至少两个模块中均过代表(图4C)。因此,CS中早期改变的lncRNA可能在调节自噬、RNA剪接和信号转导途径中发挥重要作用。lncRNA对自噬的调控参与了群居化的初期和中期,而对剪接体和磷酸肌醇代谢的调控则参与了整个过程。与神经递质释放有关的突触小泡循环途径的lncRNA仅在模式a(早期改变)模块中被过代表(图4C)。该结果表明,早期改变的lncRNA可能在CS期间调节神经系统中具有重要作用。此外,一些lncRNA与已知的型变相关基因密切相关。例如,早期改变的lncRNALNC531328.2、LNC1425451.2、LNC1088763.7和LNC492755.1与基因NPYR、NPF1a和Vat1相关(图4A)。持续变化的lncRNALNC494161.1和LNC1065048.14与多巴胺途径中的基因Ebony和Vat1相关。在CS网络中,程度值前5%的lncRNA被视为hublncRNA(图4A)。计算了lncRNA基因座的程度值,在CS中,基于度数排名前5%的10个lncRNA基因座被鉴定为hublncRNA。其中,四个基因表达上调,其他六个基因表达下调(图4D)。在IG期间,包括谷氨酸能突触、多巴胺能突触和胆碱能突触途径在内的与突触有关的途径得以丰富。这些途径中的大多数都富含早期改变的模块。同时,多巴胺能突触和胆碱能突触途径仅参与模式a(早期改变)的模块(图4E)。IG中也丰富了信号转导途径,例如钙信号传导、Ras信号传导、胰岛素信号传导和MAPK信号传导途径。功能注释的结果表明,与CS相比,IG中早期变化的lncRNA参与的突触相关和信号处理途径更多。
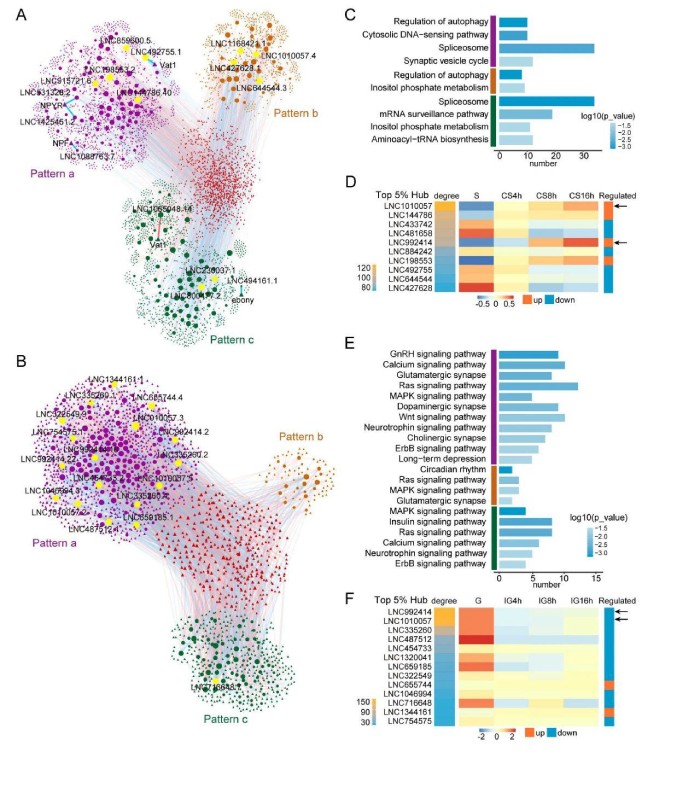
图4早期改变的lncRNA参与CS和IG的不同途径
5、LNC1010057可能调节蝗虫型变
为为了验证LNC1010057和LNC992414在蝗虫型变中的功能,进行了一系列分子生物学实验。首先,克隆了在LNC1010057和LNC992414基因座中鉴定出的长的转录本。LNC1010057由重复的A、B和C元素组成,这些元素依次分布并重复4次,但C元素重复三次(图5A)。序列比对证明LNC992414在5’端与LNC1010057共享相似的重复序列,但是它们是从不同的基因组基因座转录而来的(图5A)。尽管LNC992414的定量表达水平可以通过特异性引物检测,但LNC1010057的表达水平为重复元件的总表达水平。如预期的那样,LNC1010057和LNC992414的表达模式极为相似。在CS期间,两种lncRNA的表达持续增加,并且在16h几乎增加了四倍(图5B)。在IG期间,4h后表达水平显着下降,之后保持相对稳定(图5B)。LNC1010057和LNC992414之间的序列结构和表达模式的相似性表明它们是同源的lncRNA。但是,实时qPCR分析表明,在散居型和群居型蝗虫的大脑中,LNC1010057的表达水平比LNC992414的表达水平高约1000倍(图5C)。其次,为了测试LNC1010057和LNC992414是否参与蝗虫型变调节,在通过RNAi抑制蝗虫大脑中的表达后进行了行为分析。显着降低LNC1010057的表达水平(7470对3764610)和LNC992414(1.9vs.1.0)(图5D)后行为分析表明,群居型蝗虫其行为显着改变为散居状态(图5E)。此外,多个与型有关的行为参数发生了变化。总移动距离(TDM)和总移动持续时间(TDMV)显着降低(图5F),但是,移动速度没有区别。同时群居蝗虫的特定喜好行为显着下降(58.3对-13.5图5F)。但LNC992414表达降低并未引起从群居状态到散居状态的转变(图5G和H)。与对照相比,与型相关的行为参数(包括运动和特定物种的优选行为)没有差异(图5I)。LNC992414的RNAi实验不会引起行为变化,因此排除了LNC992414调节蝗虫型变的可能性。这些结果表明是LNC1010057而不是LNC992414潜在地调节了蝗虫的型变。
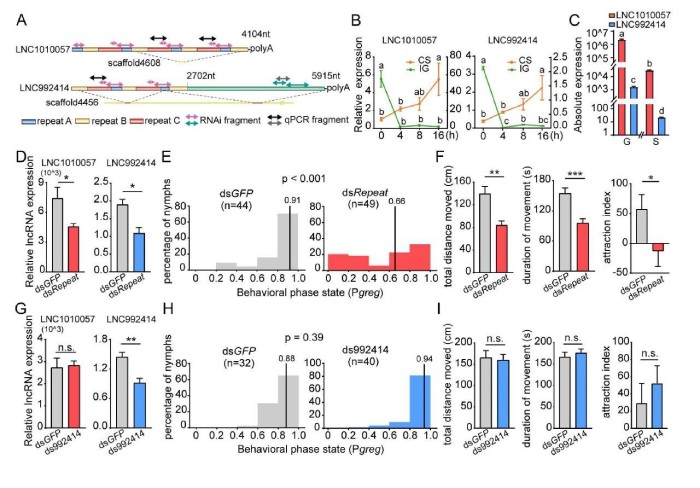
图5LNC1010057潜在地调节了蝗虫的型变
总结
lncRNA被证实是生物过程的关键调控因子,调节包括mRNA转录、稳定性、翻译和翻译后修饰等多种生物途径。之前研究表明昆虫lncRNA参与了例如杀虫剂抗性、繁殖力和腺体凋亡等生物过程,但是尚未有证据证明lncRNA可以调节非模型昆虫的行为。蝗虫是世界范围内的一种农业害虫,表现出显着的表型可塑性。为了鉴定与其型变相关的lncRNA,本文系统地分析了蝗虫lncRNA的表达并注释其功能,证实与mRNAs相比lncRNAs显示对型变更敏感的响应,并证实了其中一个lncRNA可调节型变相关行为。本研究揭示了lncRNA在蝗虫型变中的重要作用以及表型中蛋白编码基因和lncRNA之间的相互作用。深入解析蝗虫散居型和群居型转变的分子机制,为可持续治理蝗虫的新策略和新方法的开发提供了基础。




 京公网安备 11011302003368号
京公网安备 11011302003368号