

英文题目:Genomic introgression through interspecific hybridization counteracts genetic bottleneck during soybean domestication
中文题目:通过种间杂交实现基因组渗入抵消了大豆驯化过程中的遗传瓶颈
发表期刊:Genome Biology
影响因子:14.028
发表时间:2019年1月30日
研究背景
已有文献证明通过自发杂交和回交,农作物及其野生近缘种之间有渗入,是遗传物质转移的证据。而对渗入的进化模式、后果及其对作物驯化、品种多样化过程的影响知之甚少。种间杂交都有什么实例?本篇文章为您解析。
大豆(Glycine max [L.] Merr.)是世界上最重要的经济作物之一,为饲料、食品及工业用途的植物油、燃料提供了高质量蛋白质来源。栽培大豆起源于中国,在距今约6000至9000年前由野生大豆(Glycine soja)驯化而来,之后进行了品种多样化,形成许多适合于不同生态区域在农业系统中种植的大豆地方品种。尽管单一来源模型已被普遍接受,但大豆的驯化历史和过程仍不清楚。一项早期研究提出,从野生大豆到驯化大豆的过渡是一个渐进过程。在中国的大豆种植区仍存在形态介于野生大豆与驯化大豆间的半野生型大豆(Glycine gracilis)。由此推测从野生大豆到栽培大豆可能存在基因流。
杂交产生的基因组片段渗入是基因流的主要途径,在玉米和水稻种均报道存在基因渗入情况,但少有研究调查作物驯化期间基因组渗入的过程、模式和进化结果。
技术路线
研究结果
全基因组鉴定的野生大豆-栽培大豆基因渗入
通过302份已公布的大豆种质进行同源遗传关系分析 (Identical By Descent,IBD), 发现绝大多数大豆均存在种间基因渗入片段。各个基因组(包括62个野生种和129个地方种)中检测到的渗入片段的比例在0.00037至0.60之间,平均为0.032(图1)。
图2显示了检测到的基因渗入在各个基因组中具有> 5%渗入片段的种质的染色体分布。在野生大豆基因组中,检测到栽培大豆片段的比例为0.00059至0.41,平均为0.019。在栽培大豆基因组中,检测到野生大豆片段比例为0.00037至0.60,平均为0.031。野生大豆和栽培大豆亚群中,基因渗入片段的43.94%和54.61%由两个或更多的种质共享,其余的是种质特异性的。渗入片段均未完全固定在野生大豆或栽培大豆中(图2)。
具有推定基因渗入区域的D统计量显着低于没有推定基因渗入区域,并且也显着低于全基因组平均水平,这表明野生大豆和栽培大豆间的基因流与这些基因区域有关(图3)。
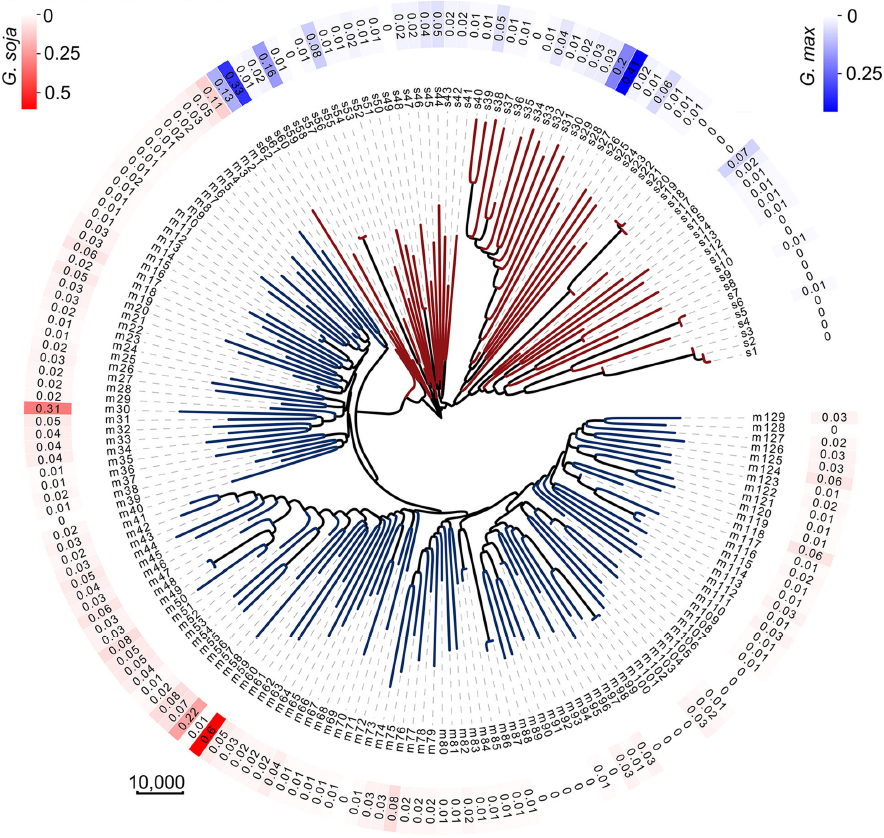
图1 野生大豆和栽培大豆的进化树
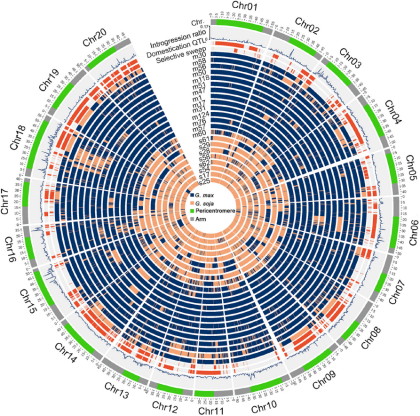
图2 种间基因渗入在全基因组的分布
环形从外至内依次表示(i)染色体臂(灰色),近着丝粒区(绿色) (ii)群体水平的基因渗入比例在各条染色体的分布(iii)驯化相关QTL位点在各条染色体的分布(红色) (iv)选择扫荡区域在各条染色体的分布(红色) (v)具有代表性的12份栽培大豆及10份野生大豆材料基因渗入区间的在各条染色体的分布:栽培大豆片段(蓝色),野生大豆片段(橙色)
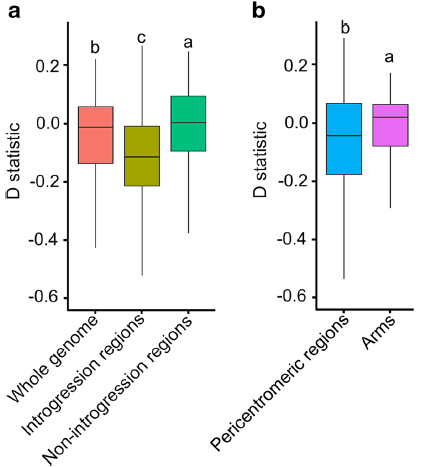
图3 D-statistic 分析方法展示野生和栽培大豆间基因组不同区域中不同模式的基因流
为了检测到基因渗入的起源,比较了野生品种(PI 578357,s61)和地方品种(PI 339734,m30)中的代表性大渗入片段,据估计分别有33%、31%的基因渗入片段具有其他种质的相应区域。图4b说明了在野生种PI 578357(s61)中,与栽培种进化相邻的野生种之一的全基因组推定性基因渗入。PI 578357中2号染色体的检测区域与地方品种黑河小黄豆(m104)中的相应区域具有最高的序列相似性(图4a,b,d,f ),而PI 578357的非渗入区与野生品种PI 522226(s5)的对应区域具有最高的相似性(图4b,c,e,f)。地方品种PI 339734中19号染色体的渗入区域与野生大豆PI 407275(s42)中的相应区域具有最高的序列相似性(图4h,i, k,l),PI 339734非渗入区与地方品种PI 548456(m111)对应区域具有最高的相似性(图4g,h,j,l)。
根据基因序列分析,PI 578357和黑河小黄豆之间及PI 339734和PI 407275之间的分歧时间分别可追溯到约0.37和27万年前(mya)。由于大豆的驯化发生在大约6000-9000年前,两对(野生大豆-栽培大豆)品种之间所检查的渗入区域的如此高的相似性应被视为野生大豆-栽培大豆渗入的直接证据。
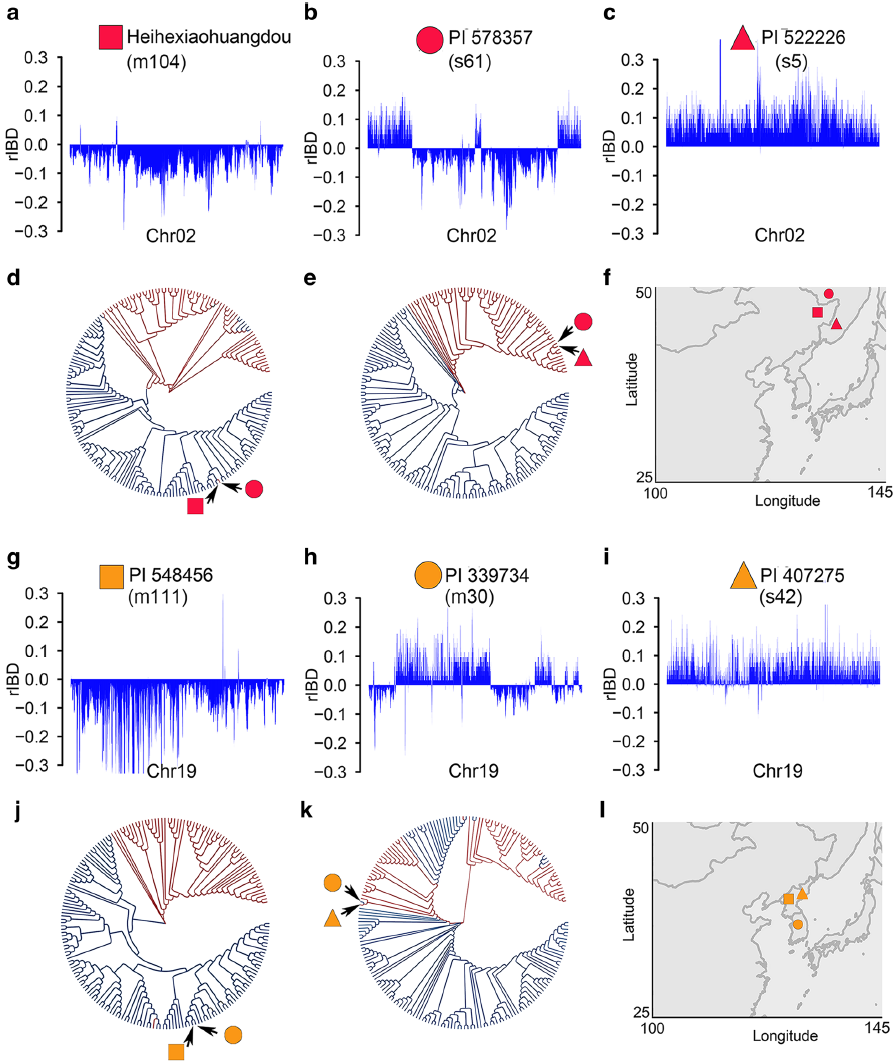
图4 基于同源遗传关系分析推测的渗入片段起源的范例
(a-c)黑河小黄豆, PI 578357和PI 52226三个材料中基因组成分在2号染色体的分布(d)基于2号染色体基因渗入区域SNP构建的进化树揭示的PI 578357中PI 52226成分的起源(e)基于2号染色体非基因渗入区域SNP构建的进化树揭示的PI 578357中黑河小黄豆成分的起源(f)上述三个材料的地理分布(g-i) PI 548456, PI 339734和PI 407275三个材料中基因组成分在19号染色体的分布(j)基于19号染色体基因渗入区域SNP构建的进化树揭示的PI 339734中PI 548546成分的起源(k)基于19号染色体非基因渗入区域SNP构建的进化树揭示的PI 339734中PI 407275成分的起源(l)上述三个材料的地理分布
野生大豆-栽培大豆基因渗入的影响因素
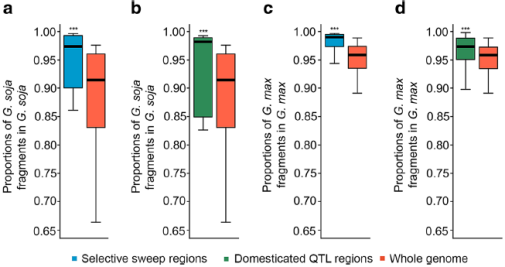
图5 自然选择与人工选择下基因渗入的模式
(a)野生大豆中选择扫荡区段中野生大豆成分的比例(b)野生大豆中驯化相关QTL区段中野生大豆成分的比例(c)栽培大豆中选择扫荡区段中栽培大豆成分的比例(d)栽培大豆中驯化相关QTL区段中栽培大豆成分的比例
相较于在野生大豆种质中检测到的基因组其他区域,与选择性扫描区域相应区域中的栽培大豆片段比例显着降低(图5a)。相比之下,与栽培大豆基因组相比,选择性扫除区中野生大豆片段在基因组其他区域的检出率显着降低(图5c)。在栽培大豆或野生大豆相应驯化QTL区域中,渗入片段比例比选择性扫描区域中检测到的小(图5b,d)。这些结果表明双向选择,即自然选择与人工选择,对野生大豆和栽培大豆中渗入片段的保留具有不同的结果、影响。
关键驯化基因周围的基因流
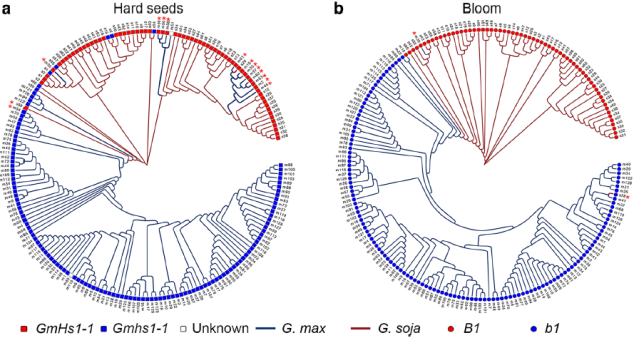
图6 基于两个驯化相关基因所在的选择扫荡区段SNP信息构建的进化树
基因渗入被认为是基因流的主要途径,那么基因流如何在种群水平上影响驯化过程和大豆基因组的遗传结构?现有研究已分离出分别控制种子硬度和种皮开花的关键大豆驯化基因GmHs1-1和Bloom1(B1),围绕GmHS1-1、Gmhs1-1和B1、b1基因座的两个选择性扫除区域中的SNP分析系统发育关系发现基因渗入现象在这两个区域均有存在。考虑到野生大豆和栽培大豆亚种的系统发育差异,在研究种群中两个驯化基因位点检测到的选择性扫描区域和单倍型的混合将被视为亚种群间基因流动的进一步证据。
核基因组和细胞器基因组之间的不对称多样化揭示了基因渗入
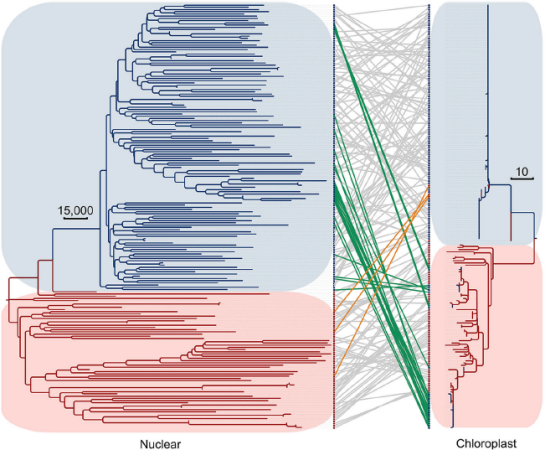
图7 栽培大豆和野生大豆核基因组和叶绿体基因组的非对称分歧
191个材料的核基因组进化树基于全基因组的SNP信息,叶绿体进化树基于333个高度可信SNP位点。栽培大豆用蓝色分枝蓝色背景表示,野生大豆用红色分枝红色背景表示。两个进化树中的相同材料用直线连接。蓝线表示栽培大豆中含有野生大豆叶绿体成分,橙线表示野生大豆中含有栽培大豆叶绿体成分。灰线表示分别具有野生大豆和栽培大豆叶绿体的种质。
栽培大豆比野生大豆的叶绿体进化分化程度更小,但有些栽培大豆中含有野生大豆的叶绿体基因组,有些野生大豆中含有栽培大豆的叶绿体基因组(图7)。说明野生大豆与栽培大豆之间存在杂交事件,这种杂交事件很可能是丰富的遗传多样性的原因。核基因组与叶绿体基因组总体上呈现出共进化趋势,但部分细胞核与叶绿体基因组间发生了非平行进化,或许是由于种内天然杂交。
选择抗渗入的后果
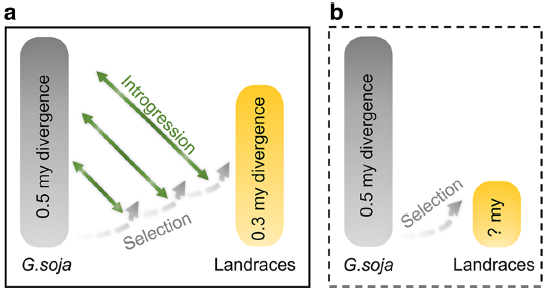
图8 大豆驯化过程模型
在大豆驯化期间或之后,种间杂交和随后的回交产生广泛的基因组渗入。之后通过从QTL或大豆驯化基础的选择清除区域清除渗入的变异,对DRT(驯化相关基因)进行重新选择或轮回选择(图8)。在这些区域中观察到的野生大豆渗入片段的比例明显低于地方品种的其余基因组区域。该模式与野生大豆种质中假定的渐渗的栽培大豆片段的分布模式相呼应,表明种间渐渗的进化命运由两个不同选择压力的相对强度决定。作物驯化伴随着遗传多样性的大量丢失,是作物形成中的主要遗传瓶颈。但与野生大豆相比,栽培大豆中依然存在着较高的遗传多样性。据此估算,栽培大豆个体间的平均分化时间可追溯到30万年前,而这样高的多样性通常是无法通过对某个地域中少数野生材料的人工选择而获得的。
结论
本研究揭示了农作物及其野生近缘种之间假定的基因组基因渗入的进化力、模式和后果,以及渗入对作物驯化和品种多样化过程的影响。设想种间渗入是抵消驯化作物特别是单种驯化作物遗传多样性减少的重要机制。






 京公网安备 11011302003368号
京公网安备 11011302003368号