
转录组分析新型四倍体水稻与育性和杂种优势特异相关的差异表达基因
Scientific Reports 2016
研究背景
水稻是最重要的粮食作物之一,在过去的50年里,通过杂交育种已经大幅度提高了水稻的产量。然而杂交育种最近几年也遇到了瓶颈,多倍体化则成为水稻育种的另一条思路。多倍体化能够提高优势基因发生重组和相互作用的可能性,提高水稻对环境的适应能力。
同源四倍体水稻(autotetraploid rice)是二倍体水稻经过染色体加倍形成的新种质。同源四倍体水稻亚种间杂种F1具有强大的生物学优势,蕴含着巨大的增产潜力,可望成为未来水稻育种的新途径。然而,其杂种F1普遍存在育性偏低,产量优势难以发挥,在农业生产上难以直接应用的问题。所以,如何创制育性正常且能克服杂种F1不育性的新型四倍体(neo-tetraploid)是利用其优势的关键。
为了避免同源四倍体水稻育性偏低的问题,华南农大某研究组经过20年的努力,培育出2个新型四倍体水稻,其结实率可达80%以上,于2016年获得国家植物新品种权。本研究旨在通过转录组测序,研究新型四倍体(Huaduo 3)与同源四倍体杂交F1代的花药、子房和叶片的基因表达情况,为解析新型四倍体育性和杂种优势的分子机制打下基础。
材料方法
新型四倍体水稻Huaduo 3(H3),40种不同的同源四倍体水稻,以及杂交F1代。同源四倍体水稻 Huajingxian 74-4x(T452)与H3的杂交F1代用于测序分析。
转录组测序:取T452,H3及杂交F1代的三种不同组织分别进行转录组测序,包括减数分裂阶段的花药、减数分裂阶段的子房、开花阶段的旗叶。每个组织每种品系有3个生物学重复,共27个样品。测序平台为Illumina HiSeq 2500,平均每个样品测约5G。
小RNA测序:取T452,H3及杂交F1代的减数分裂阶段的花药进行小RNA测序,无生物学重复。
全基因组重测序:两个亲本T452和H3的叶片DNA用于全基因组重测序。
嗯,测序手段还是挺多滴!
技术路线
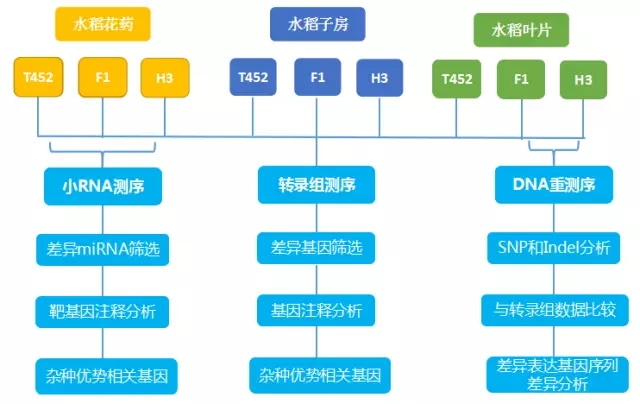
研究结果
1.新型四倍体水稻Huaduo 3(H3)的育种及其与其它四倍体水稻的杂交过程
将两种同源四倍体水稻(Jackson-4x和96025)进行杂交,获得F1代杂交种。F1代杂交种再进行连续自交,到F5代时发现一株水稻有80%的结实率,这株水稻经过多代连续自交,到F10-F13代时出现可以稳定遗传的高结实率性状,这个品系被命名为Huaduo 3(H3)。H3不仅结实率高,还有其它高产性状(表1),其花粉母细胞有90%以上是正常的。
将H3与40种其它品系的同源四倍体水稻(26个印度种和14个日本种)进行杂交,获得的40种F1代表现出了很多大大优于亲本的性状,尤其是单株谷粒数、结实率、单株产量等性状。为了进一步研究杂交F1代的杂种优势,我们挑选了T452和H3的杂交F1代进行转录组研究。T452是一种育性较低的同源四倍体,杂交F1代的高亲优势几乎都是正的,除了谷粒长度和10谷粒宽度两个指标。杂交F1代的中亲优势除了10谷粒宽度这一个指标以外都是正的,说明杂交F1代表现出了明显的杂种优势。
高亲优势(High-parent heterosis,HPH)是指杂交F1代的产量或者某一数量性状的数值与高值亲本(HP)同一性状数值差值的比率。高亲优势(%)=(F1-HP)/HP*100%。
中亲优势(Mid-parent heterosis,MPH)指杂交F1代的产量或者某一数量性状的数值与双亲(P1和P2)同一性状的平均值差值的比率。中亲优势(%)=[F1-(P1+P2)/2]/(P1+P2)/2*100%。
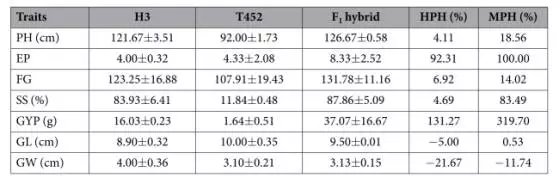
表一:T452与H3杂交F1代的杂种优势。HPH:高亲优势;MPH中亲优势;PH:植株高度;EP:单株谷粒数;FG:单花谷粒数;SS:结实率;GYP:单株产量;GL:10谷粒长度;GW:10谷粒宽度。

图1:亲本和杂交F1代的表型。(A)Jackson-4x(T45),Huaduo 3(H3)与96025(T44)的谷粒大小,H3是来自T45和T44的杂交;(B)H3在大田中的长势;(C)T45,H3,T44的植物表型;(D)Shennong15-4x(T424),H3,以及两者杂交F1代的表型。
2.转录组测序
因为T452和H3的杂交F1代表现出了优良性状,因此可以通过转录组测序研究性状差异的原因。取T452,H3及杂交F1代的三种不同组织分别进行转录组测序,包括减数分裂阶段的花药、减数分裂阶段的子房、开花阶段的旗叶。总共测了548M clean reads,平均有89.06%的reads可以比对到参考基因组上,其中93.45%都是比对到唯一位置上。三个生物学重复之间的相关性都达到了0.8以上。用qRT-PCR的方法对12个差异表达基因(DEG)的表达量进行了验证,结果与转录组测序的结果一致。对不同样品进行层次聚类分析表明,相同组织的样品总能聚到一块(图2)。
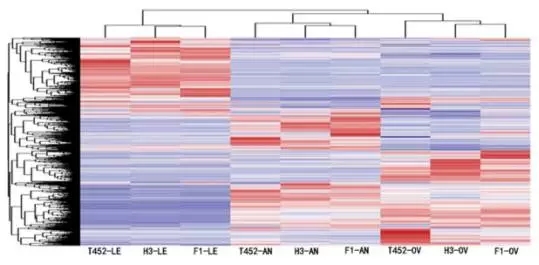
图2:所有基因的层次聚类分析。AN:花药;OV:子房;LE:叶片。
3.差异表达基因(DEG)分析及注释
在三个不同株系中共发现17877个DEG,两两比较的DEG数目在1521到2498之间(表2)。F1和亲本之间的DEG称为DEGF,亲本之间的DEG称为DEGP。DEGF的基因能分为两个不同的组,其中一个组在DEGP里面也有,另一个组只属于DEGF,后者被称为DEGFu。这些DEGFu基因能够解释F1和亲本之间的表型差异,因此接下来的研究种重点关注DEGFu基因。在这17877个DEG里面,有1150,1014,1122个DEGFu与花药、子房、叶片有关,其中分别有807,663,866个基因与花药、子房、叶片特异相关,这些基因被称为DEGFu-sp。
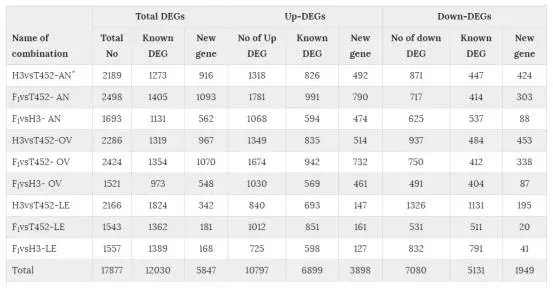
表2:三个不同组织中的差异表达基因统计。AN:花药;OV:子房;LE:叶片。
为了研究DEGFu-sp基因的功能,我们首先研究了其中的转录因子,共发现了44个转录因子,花药里面16个,子房里面10个,叶片里面18个。对DEGFu-sp基因的蛋白互作网络分析表明这些基因相互之间有显著的联系,多个代谢通路富集程度较高。
GO富集分析表明花药中的DEGFu-sp基因有19个生物学过程类别显著富集,包括光合作用、代谢途径、合成调控和转录调控。6个通路类别显著富集,包括光合作用和代谢通路。13个基因在F1中的表达量显著高于T452。在807个花药特异的DEGFu-sp中,15个基因与46个其它重要基因共表达。
接下来用同样的方法对子房和叶片中的DEGFu-sp基因进行了GO富集分析。在叶片当中有5个基因在F1中表达量显著高于T452。H3水稻叶片在开花过程中颜色逐渐变化,而T452叶片在开花过程中颜色保持不变。因此,我们比较了叶片中F1比T452上调的基因,以及H3比T452上调的基因,两者有74.01%的重合。有趣的是,在叶片中发现了两个主要的功能基因类别,分别是程序性细胞死亡和防御反应。
4. 花药特异性差异表达基因与减数分裂有关
因为T452育性较低,而F1代育性较高,为了研究育性的差异,我们重点研究了F1花药中与T452相比上调的基因。首先,有643个基因在F1花药中与T452相比特异上调,这其中排除了H3与T452相比上调的基因。对这643个基因进行GO分析表明,有6个功能基因类别富集,这些基因与防御反应、光合作用、细胞凋亡和程序性细胞死亡有关。这其中有41个基因在育性较低的同源四倍体中表达量低于二倍体。在这41个基因中,4个基因,LOC_Os04g33830,LOC_Os12g08770,LOC_Os06g21590,LOC_Os07g25430,与光合作用有关。
接下来,将这643个特异上调的基因与野生型水稻花药减数分裂相关基因的表达量相互比较,发现有9个基因都在减数分裂前期到四分体期表达。
随后,我们比较了花药特异性差异表达基因DEGFu-sp和野生型水稻减数分裂相关基因表达数据,共发现有42个减数分裂特异性基因和8个减数分裂相关基因(图3)。在8个减数分裂相关的基因中,有两个基因和DNA修复和染色体结构有关。
因为转录组也会被表观遗传调控,我们也进了小RNA测序。在花药中发现了288个差异表达的miRNA(DER),其中有13个为新发现的。在这288个DER中,有38个只在F1与亲本相比中特有,这38个基因被称为DERFu。这38个DERFu有397个靶标基因。GO分析表明这397个靶标基因有7个功能基因类别富集,这些基因与细胞凋亡、防御反应、程序性细胞死亡、细胞自噬、DNA完整性和胁迫反应有关。而且,大部分靶标基因同于F1或者H3中上调的基因。然后我们比较了这38个DERFu与之前发现的差异表达miRNA数据,发现其中一个miRNA,osa-miR5072_L-2_1ss3AG,在
Taichung65-2x和 Taichung65-4x水稻中的四倍体中下调。这个miRNA靶标基因为 LOC_Os02g24960,是一个逆转录转座子基因。
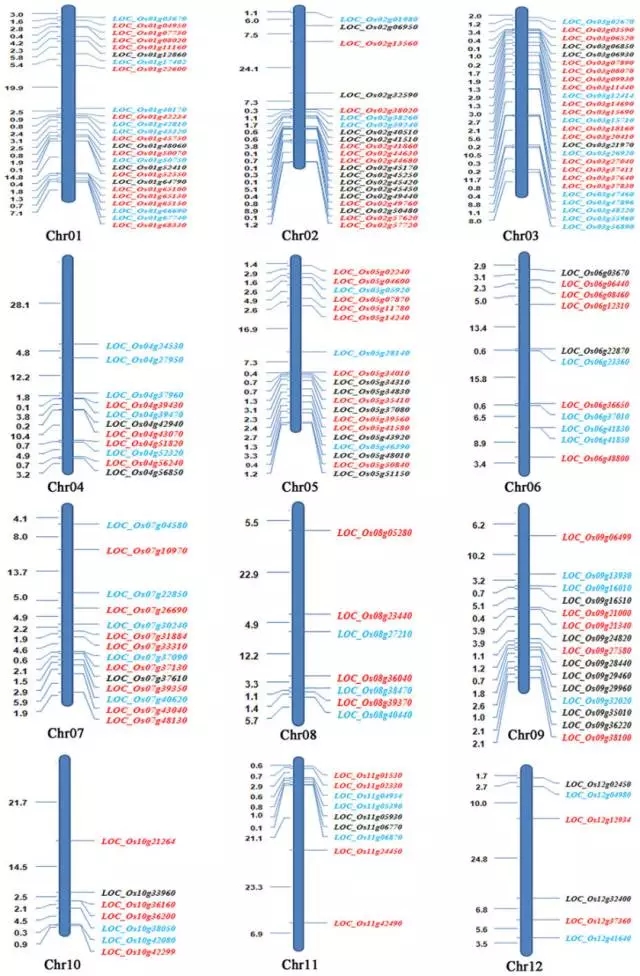
图3:与育性和杂种优势相关的基因在染色体上分布。红色为叶片中的基因,蓝色为花药中的基因,黑色为子房中的基因。
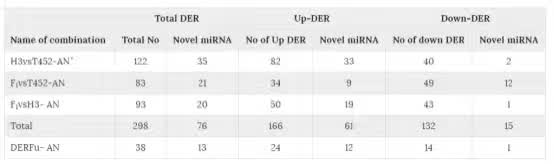
表3:花药中的差异表达miRNA(DER)列表
5.杂交F1代非累加基因表达
F1代表达的基因可以分成两个类别,一类是累加基因,一类是非累加基因。累加基因的表达量来自两个亲本的等位基因之和,非累加基因的表达量由两个亲本等位基因的平均值决定。在三个组织中共发现了1224个非累加基因(NDEG),在花药、子房和叶片中分别为314个,578个,332个。其中895个上调,329个下调。通过对花药NDEG和水稻花药减数分裂特异性基因表达数据的比较,发现了53个共有的基因。其中有7个基因在四倍体中表达量低于二倍体水稻。
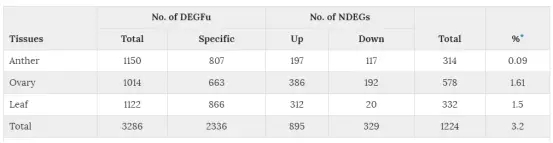
表4:非累加表达基因(NDEG)和DEGFu及不同组织的DEGFu-sp数量统计。星号表示NDEG在F1表达的总基因中的比例。
6.DNA序列差异与杂交F1代的差异表达基因之间的关系
DNA重测序表明T452和H3有很多序列差异,两者共有2351039个SNP,其中1192412个SNP在T452中特异存在,374460个SNP在H3中特异存在。两者共有499448个Indel,其中248194个Indel在T452中特异存在,84196个Indel在H3中特异存在。将T452和H3相比,SNP和Indel多态性分别为66.77%和69.97%。在这些多态性位点中,约65%的SNP和72%的Indel在基因区。47.67%的SNP和53.37%的Indel位于差异表达基因的调控区域。另外,有58908个SNP和5561个Indel位于基因编码区。
因为T452和H3存在大量的DNA序列差异,我们分析了DEGFu-sp中基因的DNA序列差异,结果表明花药、子房和叶片中分别有330个、291个、459个基因存在DNA序列差异(SNP或Indel)。花药相关的330个基因可以分为15个不同的生物学过程类别,主要和光合作用、细胞代谢、转录和生物合成有关。子房相关的基因有1个生物学过程富集,即代谢通路。同样的,叶片相关的基因有8个生物学过程富集,包括氮代谢、细胞运输等。这些富集的生物学过程基本上与DEGFu-sp富集的生物学过程一致。
另外,非累加差异表达基因中也有SNP和Indel,花药、子房和叶片中分别有67个、133个、87个基因存在DNA序列差异,并对这些基因进行了富集分析。其中25个基因为转录因子。
结论
本研究培育了一个新型四倍体水稻H3,H3与同源四倍体T452杂交产生的F1代具有育性高、产量高等优良的杂种优势性状。通过比较H3,T452,以及杂交F1代的三个不同组织(花药、子房、叶片)的转录组差异,尤其是重点分析了花药的转录组差异,找到了多个与减数分裂有关的差异表达基因,这些基因可能与杂种优势有关。对花药的小RNA测序也找到了一些F1中差异表达的miRNA,这些miRNA的靶基因也与转录组测序中上调的基因有大部分重合,说明miRNA可能参与调控了杂交F1代的性状差异。对两个亲本DNA序列的差异进行全基因组重测序分析,并与转录组数据进行联合分析,找到了DNA层面上两者育性差异的可能原因。
创新点
培育出高结实率的新型四倍体水稻,可以用于四倍体水稻育种;
转录组测序比较了新型四倍体水稻,同源四倍体水稻及杂交F1代不同组织的转录组差异,找到了影响四倍体水稻育性和杂种优势的相关基因;
小RNA测序和全基因组重测序结果与转录组数据相结合,深入解析性状差异原因。
点评
这篇文章选取的实验材料很好,性状优良,而且研究得很深入,比较了三种水稻三个不同组织的转录组,还进行了小RNA测序和全基因组重测序。通过深入分析,找到了一些影响水稻育性和杂种优势的相关基因,为后续的育种工作打下了基础。
对多种测序手段结合分析感兴趣的小伙伴快来试试吧!
参考文献
Guo H, Mendrikahy J N, Lei X, et al. Transcriptome analysis of neo-tetraploid rice reveals specific differential gene expressions associated with fertility and heterosis[J]. Scientific reports, 2017, 7:40139.




 京公网安备 11011302003368号
京公网安备 11011302003368号