

2024年7月,中国医科大学盛京医院在国际学术期刊cell proliferation发表一项重要研究成果,题为:Rack1‐mediated ferroptosis affects hindgut development in rats with anorectal malformations: Spatial transcriptome insights。使用空间转录组测序、免疫荧光、CCK-8、WB、ROS检测等来剖析直肠肛门畸形的发育机制。

期刊名称:cell proliferation.
影响因子:5.9
合作单位:中国医科大学盛京医院
研究部位:大鼠胚胎
研究方法:空间转录组、免疫荧光、CCK-8、WB、ROS检测等
百迈客生物为该研究提供了空间转录组测序服务。
研究背景
肛门直肠畸形 (ARM) 是儿童常见的先天性消化道畸形,发病率为 1/5000。ARM 的病因仍然难以捉摸,其发病机制与遗传和环境因素有关。最近的研究强调了遗传调控在这一过程中的关键作用。ARM 通常发生在妊娠 4-8 周,主要在出生后诊断,通常需要手术干预作为治疗。然而,ARM 患者的长期术后结局并不理想,通常会导致并发症,例如大便失禁和慢性便秘。这些并发症对受影响儿童的生活质量和社会心理发展有重大影响。作者的空间转录组学分析确定了铁死亡的发生,这是一种铁依赖性的程序性细胞死亡,其特征是活性氧 (ROS) 和脂质过氧化产物在 ARM 后肠内积累。作者假设这个过程受活化 C 激酶受体 1 (Rack1) 的调节,细胞质蛋白与鸟嘌呤核苷酸结合蛋白有相似之处。它在细胞生长、分化、信号传导和免疫反应中发挥着多种作用,并且在胚胎神经发育中具有潜在的意义。然而,以前的研究没有报道 Rack1 在铁死亡调节中的作用。因此,它在胚胎消化道发育和 ARM 形成中的作用需要进一步探索。
材料方法
使用Wistar 大鼠,在 ARM 组中,大鼠在 GD 10开始口服 1% ETU 125 mL/kg,对照组接受等剂量生理盐水。从 GDs 14-16取胚胎。
1、空间转录组测序剖宫产后,立即将取出的胚胎置于冷生理盐水中以去除表面血。将胚胎以水平矢状方向放置在含有预冷OCT的包埋盒中储存在 -80°C 冰箱中。共计6份样品,切片包括尿道和后肠层。2、免疫荧光
选择清晰全面显示尿道、后肠、URS 和尿道回瘘的石蜡切片进行染色。
研究结果
1.空间转录组测序的注释聚类
在这项研究中,作者在 GDs 14-16 (称为 N14-16) 上使用正常的 Wistar 大鼠胚胎,在 GDs 14-16 (称为 A14-16) 上使用 ETU 诱导的 ARM 胚胎进行空间转录组测序。制备大鼠胚胎的 6 个冷冻中矢状切片,每个 10 μm 厚。这些切片包括尿道和后肠层,提供了特定时间点泄殖腔发育的全面视图(图 1A)。使用 10× Genomics Visium 空间转录组技术,然后进行组织透化、cDNA 合成和文库构建,作者成功地捕获了正常和 ARM 胚胎中的视觉基因转录数据,主要来自泄殖腔区域。为了确保来自同一区域的各种切片样本之间的可比性和一致性,作者进行了聚类注释,产生了七个不同的聚类。这些集群包括泄殖腔区域内的五个特定区域(集群 0-4),即尿道、后肠、膀胱、URS 和生殖器结节。此外,两个簇 (簇 5 和 6) 对应于椎体和神经管区域 (图 1A)。平均而言,每个解剖区域覆盖 3005 个点,每个点捕获 16,905 个基因(图 1B)。
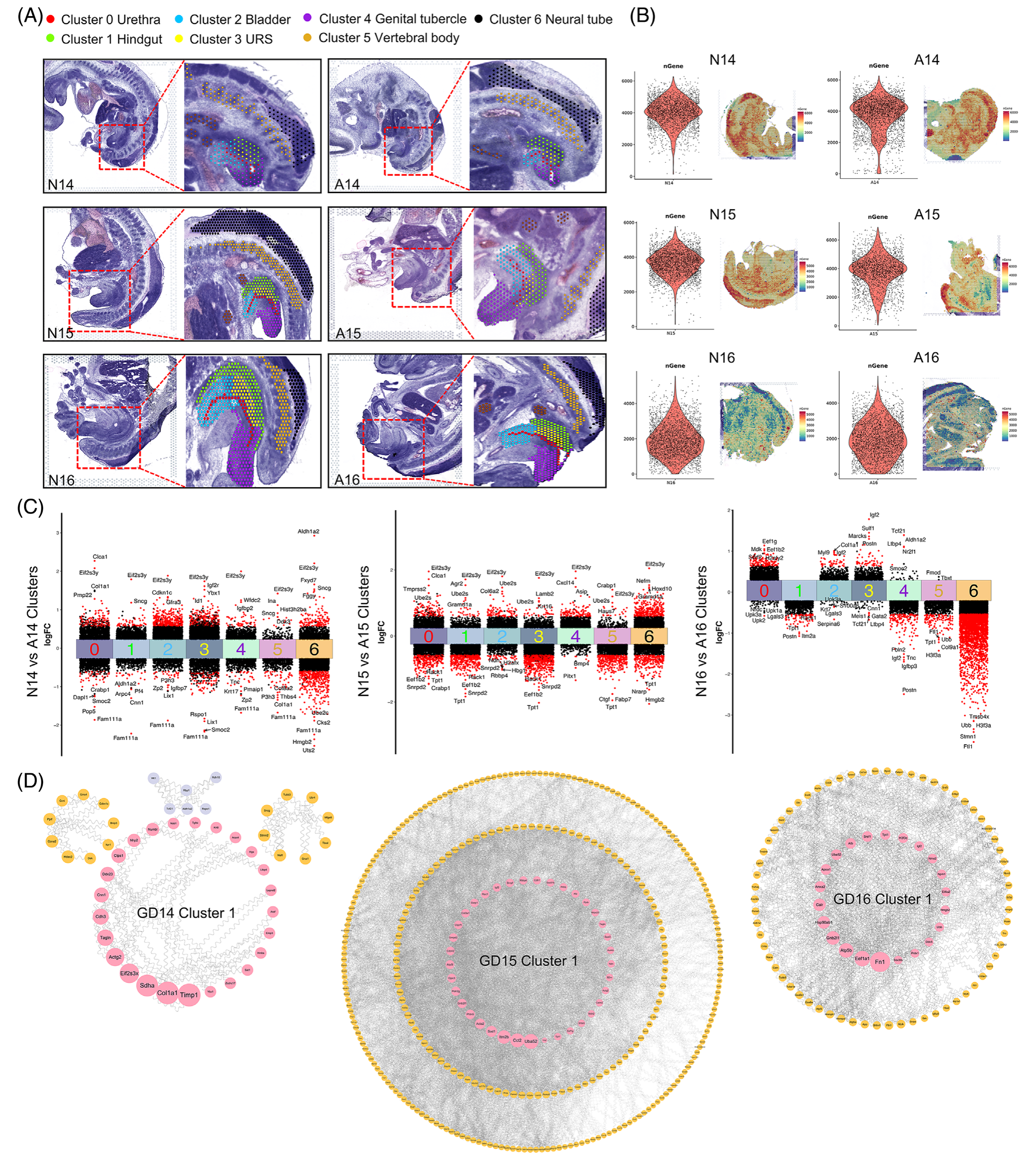
图1-空间转录组测序的注释聚类
2.正常组和 ARM 组之间的 DEG 筛选
鉴定 DEG 的标准被确定为绝对对数2 倍变化 (|log2FC|) ≥ 0.58 且显著性水平为 p< 0.05. 这种筛选导致了差异火山图的创建,以说明基因表达的改变。在 GDs 14、15 和 16 的后肠发育背景下,作者分别鉴定了 91、439 和 119 个基因,它们在这些时间点表现出不同的表达谱(图 1C)。值得注意的是,Rack1 (Gnb2l1) 在 GDs 15 和 16 上在 ARM 组后肠内的表达降低。为了进一步探索潜在的分子相互作用,作者进行了全面的 PPI 分析。通过利用 BC 值作为排名标准,作者突出了最显着的差异表达后肠基因(图 1D)。枢纽基因,在环状排列中用粉红色节点表示,包括 Uba52、Rack1 和 Tpt1,在后肠发育的 GDs 15 和 16 期间,每个基因在基因网络中都具有显着的中心性。值得注意的是,Rack1 表现出卓越的连通性,在 GD 15 和 16 的 PPI 网络中跻身前五名基因之列。
3.在 GD 15 上验证后肠部分 DEGs
图 2A-E 全面说明了 GD15 截面泄殖腔区域中前五个基因的蛋白质表达模式和定位:Rack1、Npm1、Eef1b2、Uba52 和 Tpt1。这些数字是使用免疫荧光染色获得的。值得注意的是,这 5 种蛋白质在 N15 泄殖腔区域的尿道和后肠上皮内均表现出位点特异性表达。除 Tpt1 外,其他 4 种蛋白在 AM 区域内表现出表达水平升高。此外,观察到这些蛋白质离散分布在 URS 的下肢和后肠的间质附近。相比之下,这五种蛋白的表达在 A15 泄殖腔区域的尿道和后肠上皮内明显下降,伴随着尿道直瘘部位的表达水平降低 (p < 0.05)。图 2F 显示了平均光密度 (AOD) 的半定量评估,特别是在后肠结构域内。
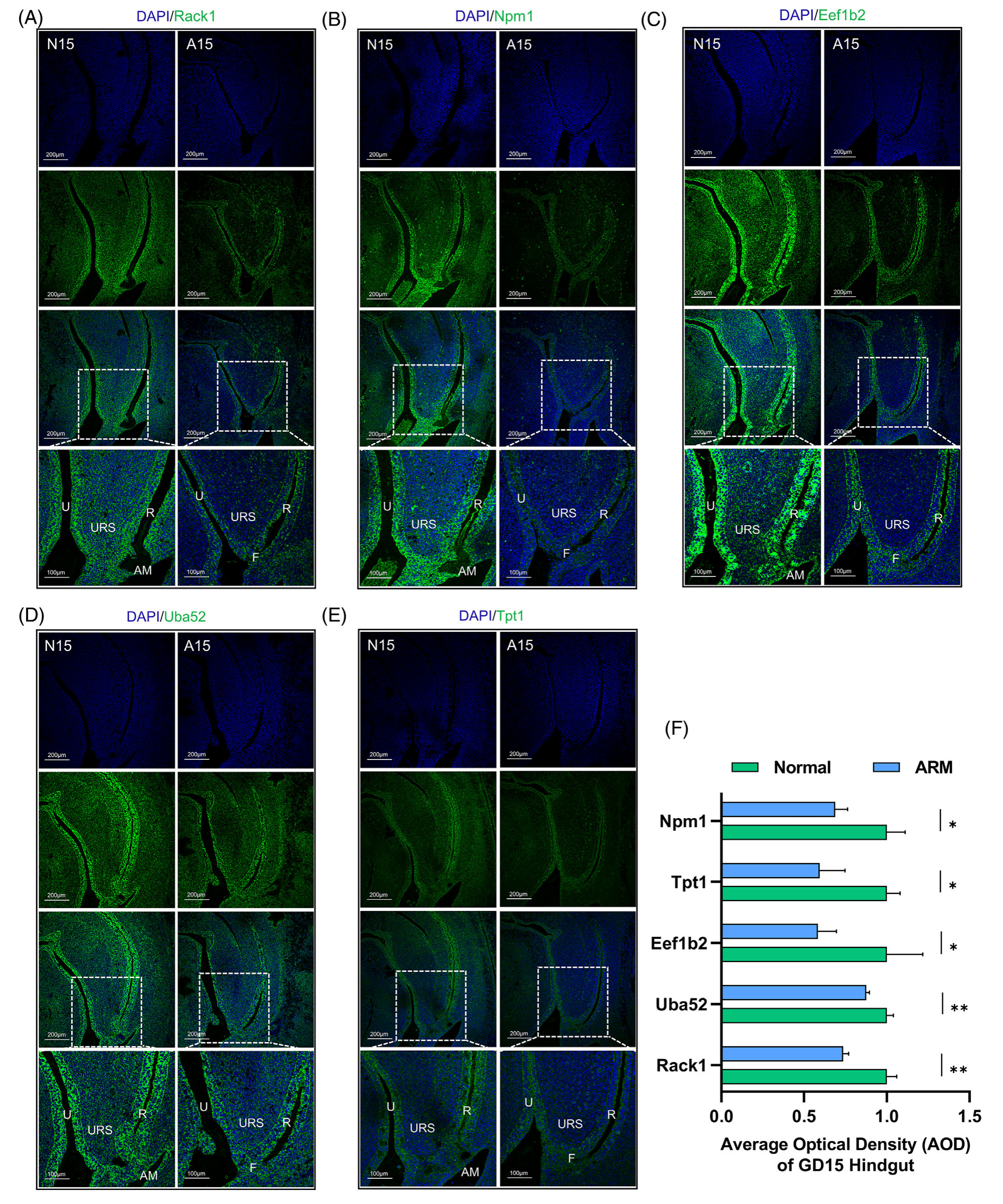
图2-在 GD 15 上验证后肠部分 DEGs
4.PROGENy 算法预测后肠中 MAPK 信号通路的集中富集
PROGENy 算法用于通过下游基因表达的变化评估信号通路的活性。作者获得了正常组和 ARM 组 GDs 14-16 样本中不同通路的活性水平评分。在使用 NNMF 对后肠区域(N14 因子 16、A14 因子 15、N15 因子 15、A15 因子 20、N16 因子 18 和 A16 因子 10)进行聚类分析中,PROGENy 算法显示与 MAPK 信号通路呈强正相关,表明 MAPK 信号在后肠中显著富集(图 3A)。从这些区域提取来自 Erk、P38、JNK 和 Erk5 家族的 12 个成员分子的定位表达模式。具体来说,Mapk3 、 Mapk6 和 Mapk14 主要富集在泄殖腔区域。这些蛋白质的免疫荧光验证证实了它们在尿道和后肠区域的特异性表达,在 URS 和间质区域的表达相对较弱,这与 PROGENy 预测一致(图 3B)。
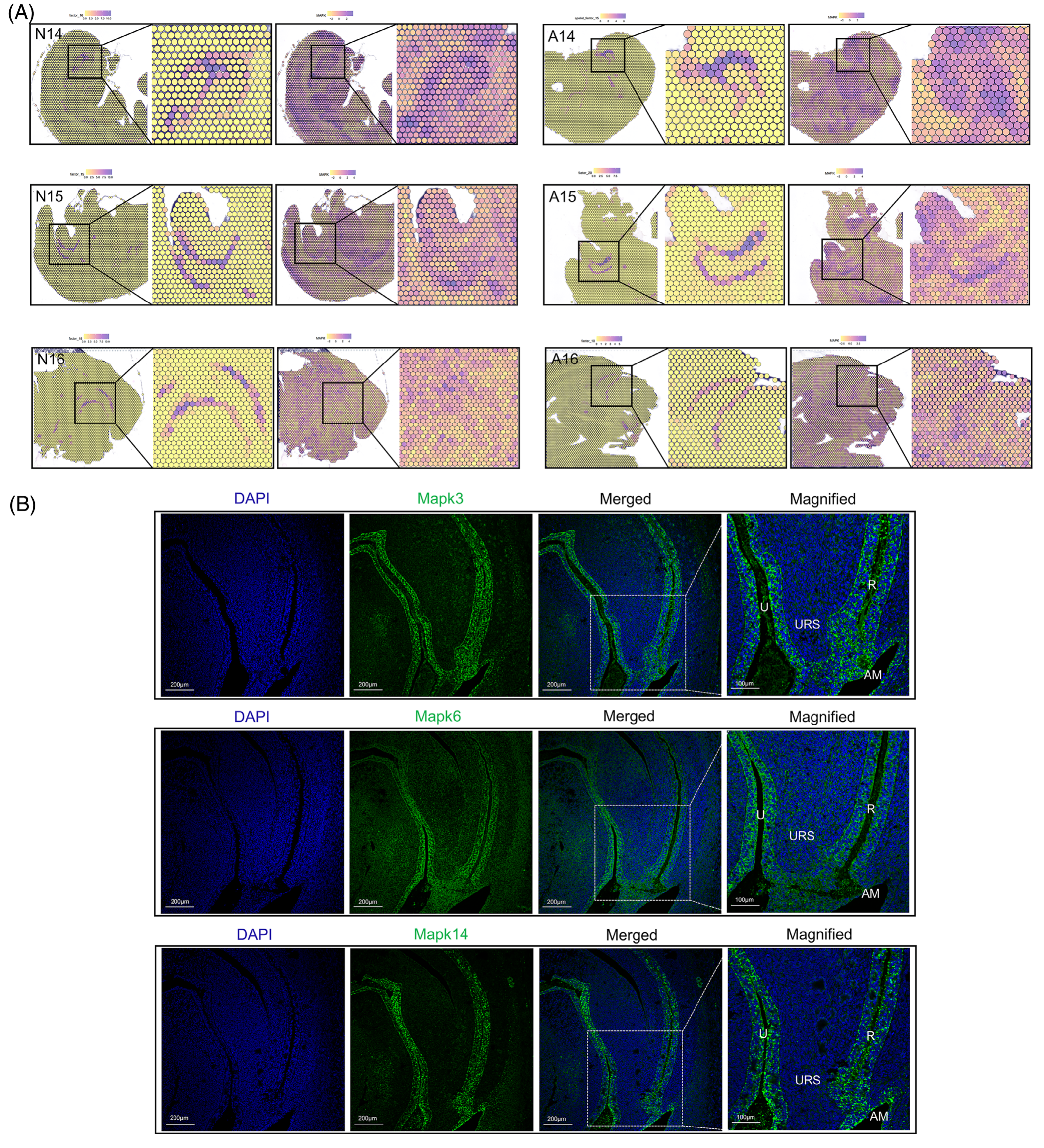
图3-PROGENy 算法预测后肠中 MAPK 信号通路的集中富集用
5.通过细胞死亡基因定位探索 Gpx4 的时空表达模式
在基因集富集分析(GSEA)数据库中,细胞死亡基因集包含161个基因,其中 7 个细胞死亡相关基因是从基因集与GD 15上簇1中的 DEGs 交集中获得的(图 4A)。圆形热图显示了与聚类1的6个样本相同区域内的细胞死亡基因的表达谱。值得注意的是,发现谷胱甘肽过氧化物酶家族的关键成员 Gpx4 在 GD 15 的簇 1 中下调(p < 0.05;图 4B)。免疫荧光染色显示 Gpx4 蛋白在尿道和后肠上皮以及 AM 区域的显着定位,在 URS 中观察到的分布相对较浅。在 ARM 组的泄殖腔中观察到 Gpx4 阳性信号的减少(图 4C)。AOD 分析表明,在 GDs 15 和 16 上,后肠中 Gpx4 表达在统计学上显着降低(p < 0.05;图 4D)。
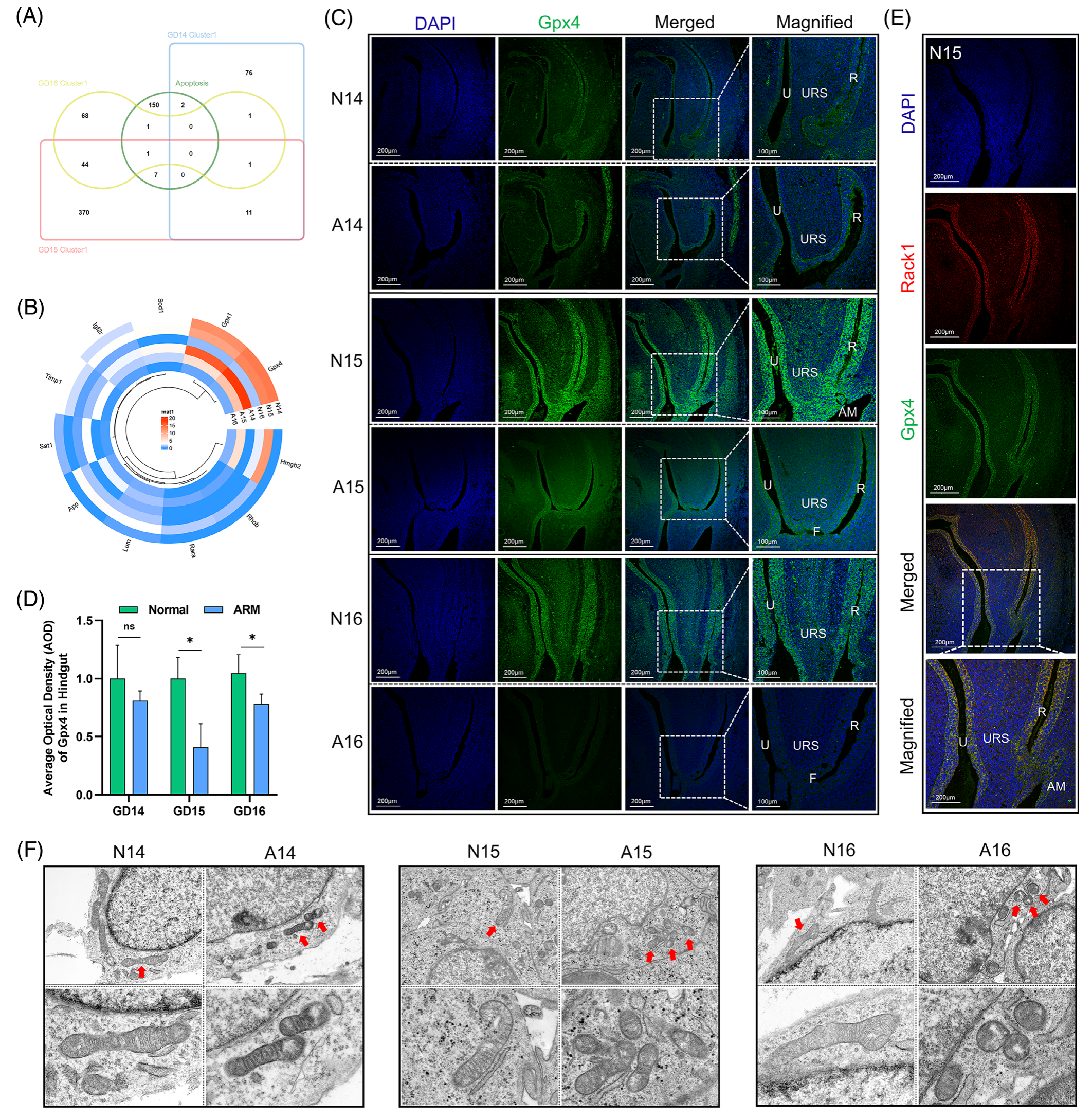
图4-通过细胞死亡基因定位探索 Gpx4 的时空表达模式
6.PD-L2 过表达抑制肿瘤细胞凋亡并促进对 EGFR-TKI 的耐药
N15 胚胎石蜡切片的双免疫荧光染色显示 Rack1 和 Gpx4 蛋白在尿道和后肠上皮内共定位。该结果表明 Rack1 在调节这些组织中的铁死亡中的潜在作用 (图 4E)。随后,进行显微解剖以获得正常和 ARM 胚胎的后肠组织。透射电子显微镜 (TEM) 显示 ARM 组细胞线粒体体积减小,线粒体密度增加,线粒体嵴减少。这些观察结果与与铁死亡发生相关的线粒体形态变化一致(图 4F)。
7.Rack1 在 GDs 14 和 16 上的时空表达谱
之前描述了 Rack1 蛋白在 GD 15 泄殖腔区的定位(图 2A),作者进一步研究了它在 GDs 14 和 16 泄殖腔区的定位。在正常胚胎 (N14) 中,Rack1 阳性信号主要在尿道和后肠的上皮细胞中观察到,后肠远端的浓度较高。这些信号也分散在 URS 和间充质细胞内。在 ARM 胚胎 (A14) 中,在 URS 尖端、后肠和尿道上皮观察到 Rack1 阳性细胞(图 5A)。比较分析显示,N14 和 A14 组之间 Rack1 阳性信号的 AOD 没有显着差异(图 5B)。在 GD 16 (N16) 的正常大鼠胚胎中,Rack1 表达在尿道和后肠上皮中仍然突出,但在 URS 和周围间充质区域受到限制(图 5A)。值得注意的是,A16 后肠区域的荧光强度降低 (p < 0.05),如图 5B 所示,这说明了后肠区域的半定量 AOD 结果。
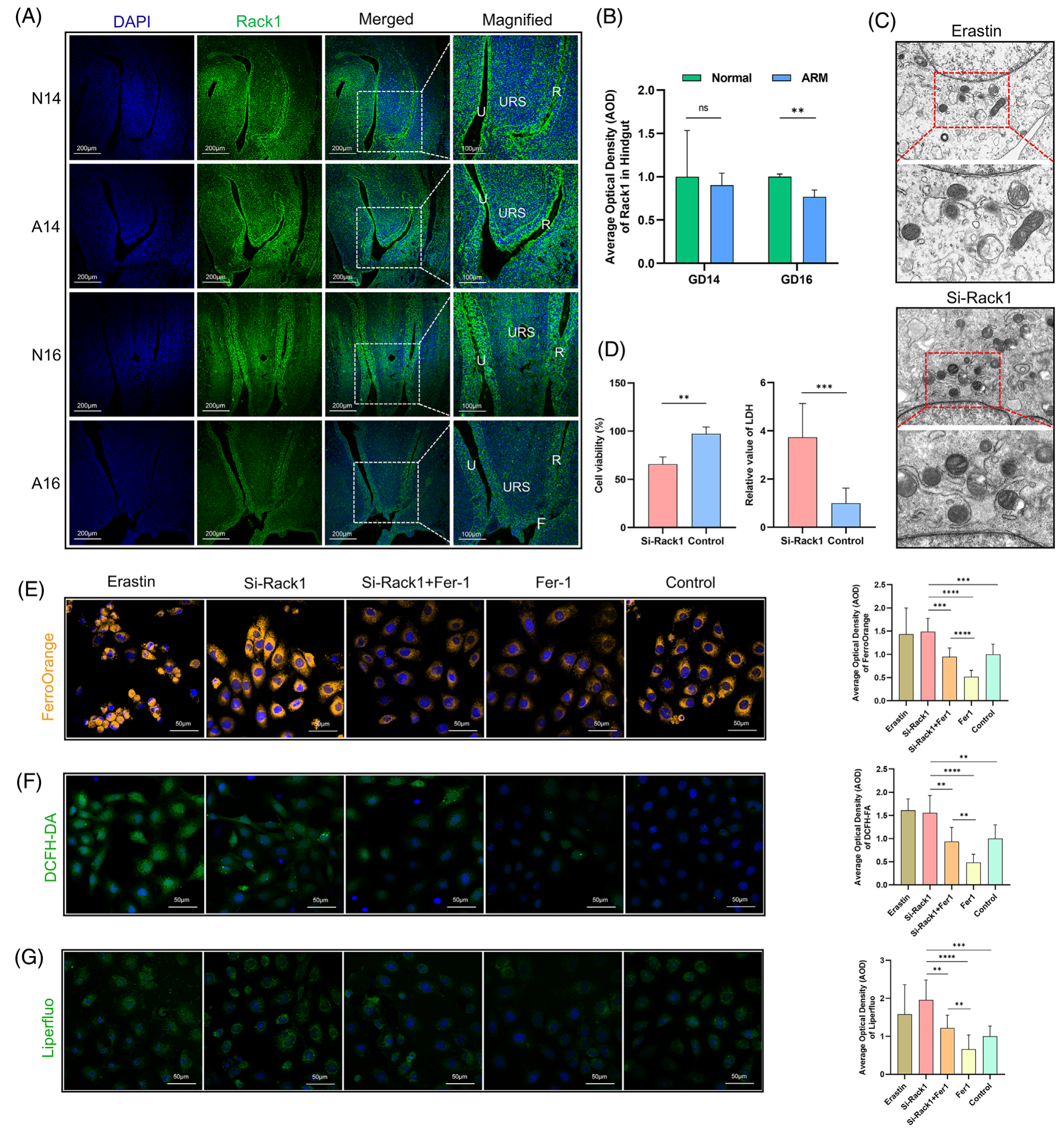
图5-Rack1 在 GDs 14 和 16 上的时空表达谱
8.敲低 Rack1 诱导肠上皮细胞铁死亡
在 IEC-6 细胞中转染 si-Rack1 后,使用 TEM 观察细胞超微结构。si-Rack1 组表现出完整的细胞和核膜,线粒体大小减小,线粒体密度增加。此外,线粒体嵴减少或不存在,类似于 erastin 处理后观察到的特征(图 5C)。如 CCK-8 测定中观察到的 Rack1 敲低导致细胞活力降低,LDH 测定所示,细胞毒性增加(p < 0.05;图 5D)。使用 FerroOrange 探针的荧光分析表明 si-Rack1 组细胞内亚铁离子浓度升高,在 Fer-1 处理后降低(p < 0.05;图 5E)。使用 DCFH-DA 探针测量细胞内 ROS 水平显示 si-Rack1 组的 ROS 水平较高,在 Fer-1 处理后下降(p < 0.05;图 5F)。Liperfluo 探针评估揭示了 Fer-1 能够减轻 si-Rack1 组中升高的脂质过氧化物水平 (p < 0.05;图 5G)。荧光强度定量结果如右图所示。
9.Rack1 敲低可增强 P38 磷酸化并调节下游 Nqo1/Gpx4 表达
如前所述,MAPK 信号通路在后肠区域显着富集(图 3A)。在 IEC-6 细胞中转染 si-Rack1 后,使用 MAPK 信号通路 PCR 阵列在 mRNA 水平上鉴定受 Rack1 影响的下游分子。使用阈值 |log2FC|≥ 2 和 p < 0.05 用于 PCR 阵列差异基因筛选,作者观察到 si-Rack1 组中 P38δ (Mapk13 encoding) 的统计学显着升高(图 6A、B)。Western blot 分析显示,Rack1 敲低后磷酸化 P38 水平增加(图 6C、D),表明对 Rack1 的干扰增强了 P38 磷酸化,从而激活了 IEC-6 细胞中的 P38-MAPK 信号通路。随后,使用铁死亡 PCR 阵列筛选 si-Rack1 和 doramapimod 处理组之间表现出 mRNA 水平变化的基因,揭示了总共 7 个基因(图 6E)。此外,这 7 个基因的表达丰度数据与现有研究报告相结合,将 Nqo1 确定为下游分子,以供进一步研究。Western blotting 分析显示,doramapimod 处理逆转了 Rack1 敲除诱导的 Nqo1 蛋白水平降低(图 6F)。此外,在 si-Rack1 组中观察到的 Gpx4 降低被 doramapimod 抑制,并通过 2-HBA 处理恢复(图 6G)。
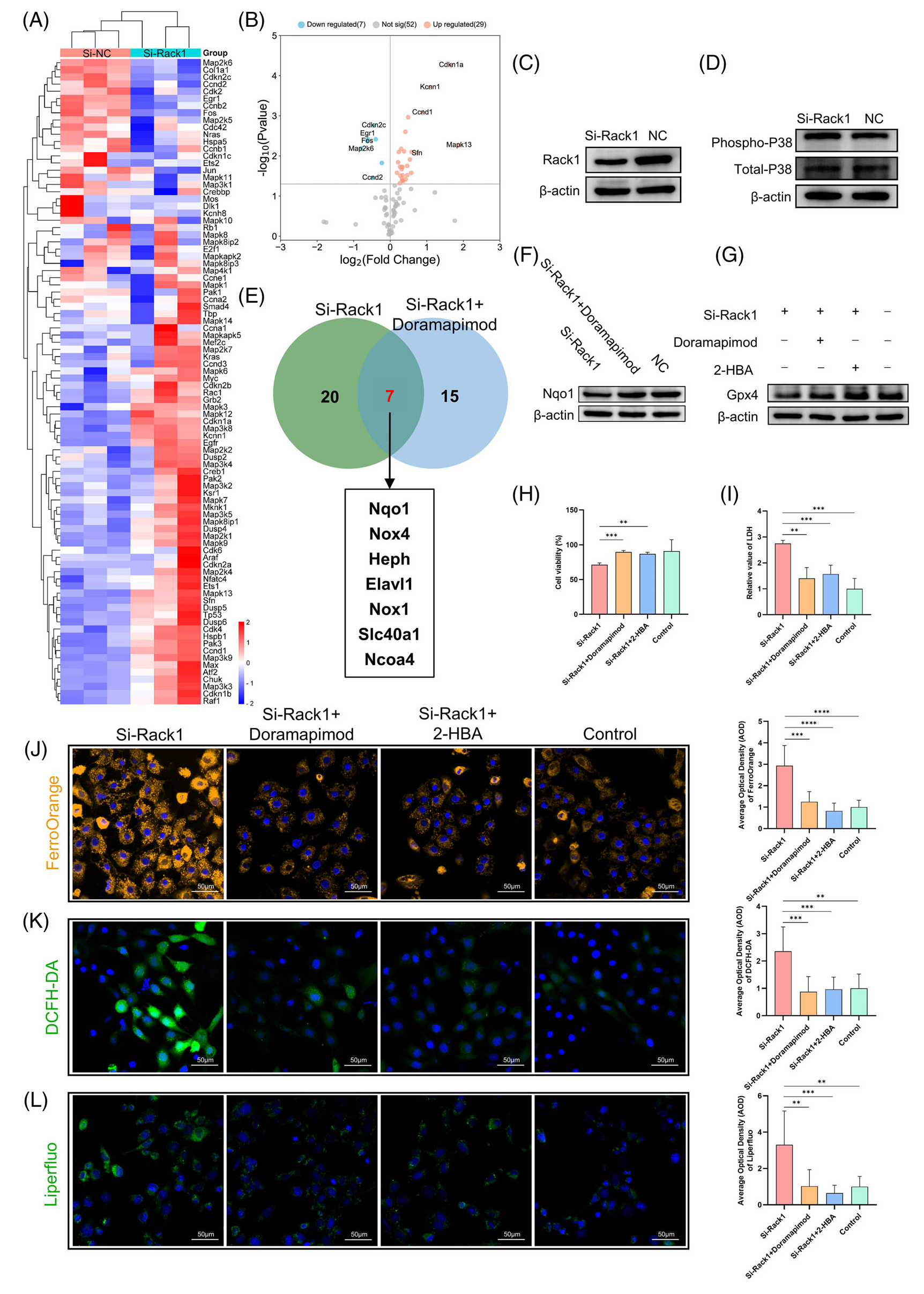
图6-Rack1 敲低可增强 P38 磷酸化并调节下游 Nqo1/Gpx4 表达
10.Rack1 通过 P38 和 Nqo1 介导肠上皮细胞中的铁死亡
在 IEC-6 细胞敲低 Rack1 的同时,使用 doramapimod 和 2-HBA 进行拯救实验。结果显示,与单独的 Rack1 敲低相比,添加 doramapimod 和 2-HBA 导致细胞活力增加和相对 LDH 含量降低 (p < 0.05;图 6H,I)。此外,在添加 doramapimod 和 2-HBA 后,细胞内亚铁离子浓度降低,同时 ROS 和脂质过氧化水平降低(p < 0.05;图 6J-L)。这些发现表明,Rack1 通过P38信号通路和 Nqo1 调节细胞内铁含量和脂质过氧化,从而介导肠上皮细胞的铁死亡。
研究总结
本研究利用空间转录组学技术对 GDs 14-16 期间的正常和 ARM 大鼠胚胎样本进行测序。作者在后肠区域鉴定了具有高连接性的新枢纽基因,即 Rack1 、 Uba52 、 Tpt1 、 Npm1 和 Eef1b2。相比之下,作者观察到 MAPK 信号通路的显着富集和 Gpx4 在后肠区域的差异表达。值得注意的是,Rack1 在 GDs 15 和 16 的 ARM 后肠中表达降低,通过 P38/Nqo1/Gpx4 轴升高细胞内铁、ROS 和脂质过氧化水平,最终诱导肠上皮细胞铁死亡,并可能影响 ARM 后肠发育。这些发现增强了作者对 ARM 发病机制的理解,并对推进 ARM 产前诊断和治疗策略的研究具有重要意义。




 京公网安备 11011302003368号
京公网安备 11011302003368号