

2024年5月9日,韶关学院和华南农业大学在国际学术期刊 Postharvest Biology and Technology 发表一项重要研究成果,题为:Metabolomics and transcriptomic profiles reveal membrane lipid metabolism being an important factor of sliced taro browning。该研究通过植物广靶代谢组和转录组分析研究了芋头片在冷藏过程中的褐变发展及潜在的褐变机制。这项研究有助于我们了解鲜切食品的褐变机制,为未来的研究和减轻冷藏条件下切片芋头褐变的潜在策略提供宝贵的见解。

文章标题:Metabolomics and transcriptomic profiles reveal membrane lipid metabolism being an important factor of sliced taro browning
期刊名称:Postharvest Biology and Technology
影响因子:7.0
合作单位:韶关学院和华南农业大学
研究对象:芋头
研究方法:生理、转录组学、代谢组学等
百迈客生物为该研究提供了植物广靶、转录组测序和分析服务。
研究背景
在饮食中加入更多的新鲜蔬菜可以带来许多健康益处,包括降低对慢性疾病的易感性和减缓衰老过程。市场上根茎类蔬菜切根产品的保质期短,破坏了这些优势。切面褐变是限制切片蔬菜产品寿命的主要因素之一。因此,了解切片产品褐变背后的机制对于开发创新技术至关重要,这些技术可以在储存和消费过程中有效地保持这些产品的营养价值和整体质量。
实验材料
本研究选用购自中国韶关的“冰琅玉”品种(Colocasia esculenta)。用于调查的芋头大小相似,从大约800克到1000克不等,没有任何明显的缺陷或机械损伤。这些选定的芋头被迅速运送到实验室,为了确保清洁,在削皮和切割之前,要用自来水冲洗掉芋头球茎表面残留的淤泥。然后,将芋头削皮,切成约1厘米厚的薄片作进一步实验。首先用次氯酸钠溶液(0.1 g/L)对芋头切片进行灭菌。随后,将无菌芋头片密封在聚乙烯袋中(0.02 mm厚,尺寸为20 × 30 cm) (Xiao et al, 2020)。最后,芋头切片冷藏(4℃), RH为90-95%,持续12天。样品每隔2天采集一次,用于后续分析。在第0天(C0)、第6天(C6)和第12天(C12)获得的样本用于植物广靶代谢组学和转录组学分析。
研究结果
1.生理分析——冷库条件下芋头片褐变评价
芋片切面颜色随贮藏时间的变化如图1A所示。切面L*值从0 d时的88.72下降到12 d时的80.36,冷藏12 d后L*值下降了9%(图1A)。芋头切片表面的a*、b*、△E和BI值在冷藏过程中也表现出类似的变化趋势(图1B-E)。这些褐变指标随着贮藏时间的延长而增加。冷藏12 d后,a*、b*、△E和BI值分别比0 d增加了582%、58%、725%和19%(图1B-E)。宏观褐变症状在第6天首次出现,并在随后的储存过程中逐渐加剧(图1F)。综上所述,这些结果表明,即使在低温(4℃)条件下,切片芋头在储存过程中也会发生褐变。
为了阐明切片芋头褐变的机制,研究人员分别在第0天、第6天和第12天对芋头样品进行了转录组学和代谢组学分析。
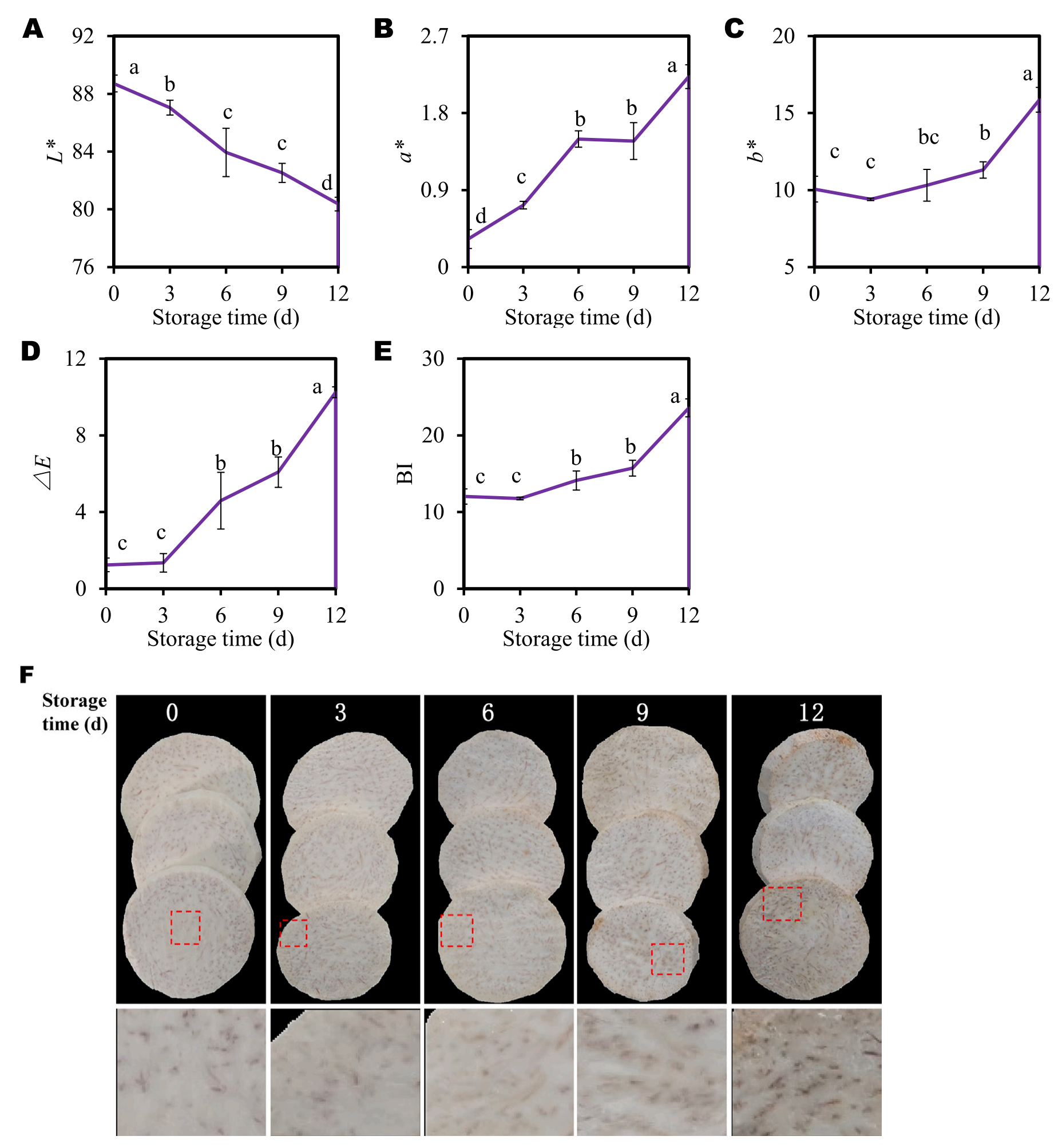
图1-4℃贮藏期间切片芋头的褐变发展
2.代谢组分析——切片芋头褐变过程代谢组学分析
为了比较芋头切片褐变过程中三个阶段代谢物组成的差异,研究通过植物广靶代谢组学分析,总共成功鉴定了芋头切片中的638种代谢物。PCA分析显示,三个储存点采集的芋头样品具有明显的分离性(图2A)。在C0 vs C6中,共鉴定出206个DAMs,其中99个DAMs的丰度增加,107个DAMs的丰度减少。C6 vs C12共197个DAMs,分别有84个DAMs丰度增加,113个DAMs丰度减少。C0 vs C12包括119个DAMs,分别有51个DAMs和68个DAMs显示丰度增加和减少(图2B)。这些结果表明,在褐变过程中,代谢物丰度的减少幅度更大。
韦恩图分析评估每个差异分组特有和共有的DAMs (图2C)。图2D-F展示了每个比较组中丰度增加和减少的top20的DAMs。C6组中10个代谢物丰度升高,如2′-脱氧腺苷、3-O-对香豆酰奎宁酸、n -乙酰- l -苏氨酸、松柏醛、9(10)-EpOME、6-O-咖啡酰丁醇、表肌醇、l -同型半胱氨酸、5-O-对香豆酰奎宁酸、异鼠李素。相反,琥珀酸酐、9-(阿拉伯糖基)次黄嘌呤、氨基丙酸、N6-(2-羟乙基)腺苷、2-(二甲氨基)鸟苷、苄基-(2 ‘-O-木糖基)葡萄糖苷、鸟苷、肌苷、D-甘露糖和肌苷5 ‘ -单磷酸在C6组中的丰度较低(图2D)。
在C6和C12中,9(10)- epOME、n -乙酰- l-苏氨酸、2 ‘ -脱氧腺苷、松木醛、2-亚麻油基甘油-1,3-二- O-葡萄糖苷、6- O -对咖啡酰基熊果甙、5-O-对香豆酰基奎宁酸、异鼠李素、反式- 4-羟基肉桂酸甲酯和3- O -对香豆酰基奎宁酸在C12组中表现出更高的富集度。相反,9-(阿拉伯糖基)次黄嘌呤、酪氨酸、S-Aiiyl-L-半胱氨酸、8,11,14-二十烷三烯酸甲酯、L-蛋氨酸甲酯、LysoPC17:0、鸟苷、肉桂酸、11、14、17-二十碳三烯酸和黄嘌呤在C6组中具有更高的丰度(图2E)。
在C0和C12中,9(10)- epOME、n -乙酰- L-苏氨酸、2 ‘ -脱氧腺苷、5- O -对香豆酰基奎宁酸、2-亚麻油基甘油-1,3-二- O -糖苷、松木醛、6- O -对咖啡酰基熊果甙、异鼠李素、3-O-对香豆酰基奎宁酸和反式-4-羟基肉桂酸甲酯在C12组中含量较高。相反,9-(阿拉伯糖基)次黄嘌呤、肌苷、S – aiiyl -L -半胱氨酸、8,11,14-二十烷三烯酸甲酯、L-蛋氨酸甲酯、LysoPC17:0、鸟苷、肉桂酸、11、14、17-二十碳三烯酸和黄嘌呤在C0组中表现出更高的丰度(图2F)。
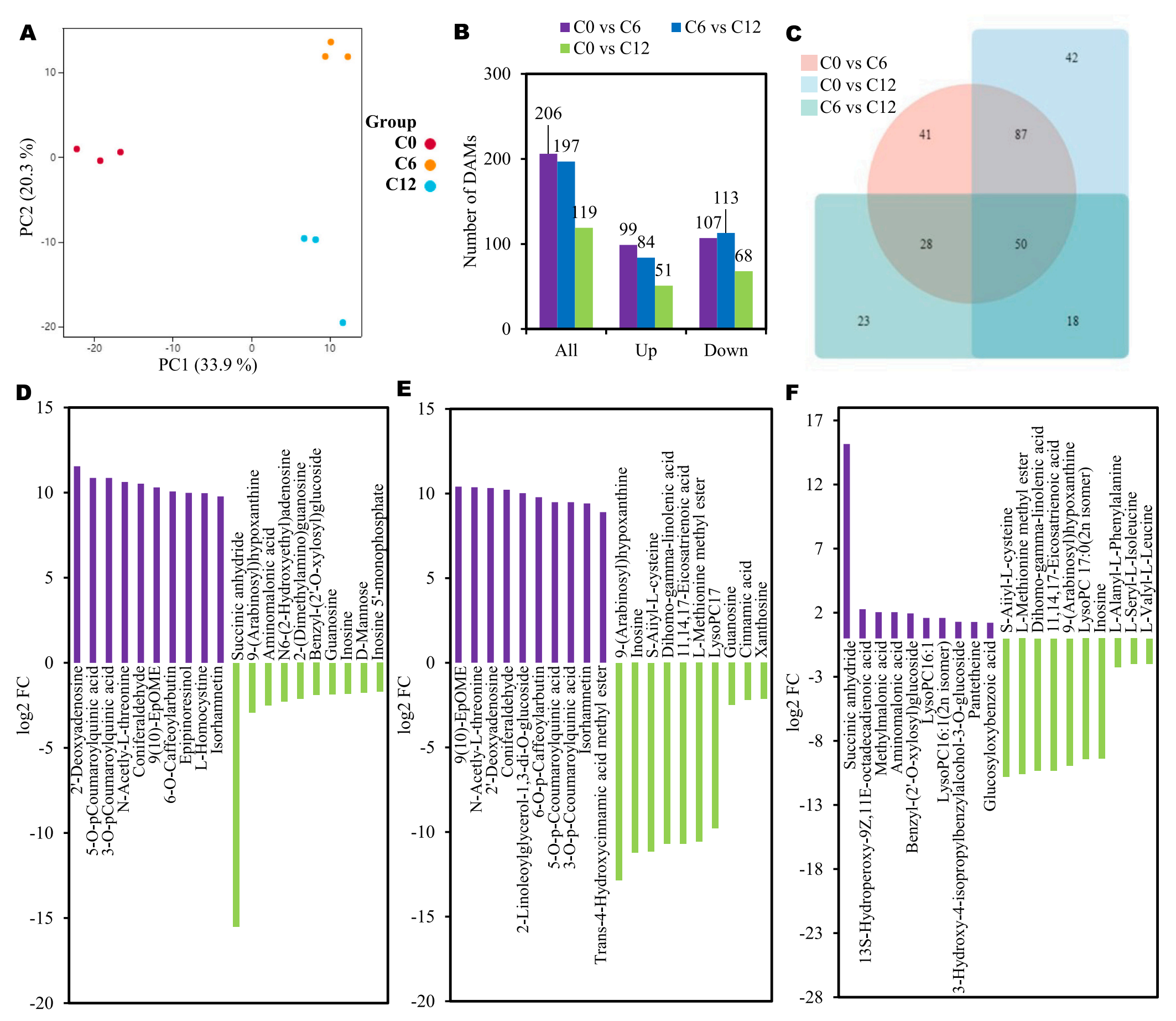
图2-切片芋头植物广靶代谢组分析
3.代谢组分析——切片芋头褐变过程中DAMs的积累模式
为进一步分析代谢物在9个样本的积累模式,该研究进行了聚类热图分析。结果显示,随着贮藏时间的延长,褐变程度逐渐增加,推测持续增加的DAMs可能有助于切片芋头的褐变或促进其褐变过程。因此,该研究关注分析在褐变过程中丰度持续上升的DAMs (图3B-L)。确定了11个DAMs在褐变过程中丰度持续增加,如2-α-亚麻烯酰甘油、甘油亚油酸、(9Z,11E)-十八烯二烯酸、N-油基乙醇胺、γ-亚麻酸、1-亚麻油基甘油、2-亚麻油基甘油、N-α-乙酰基- L-鸟氨酸、α-亚麻酸、9-羟基-10、12-十八烯二烯酸和1-α-亚麻烯酰甘油。值得注意的是,在这11个DAMs中,有10个是脂肪酸或脂质衍生物。几种褐变指标与这些水DAMs之间的相关系数非常高。这些结果表明,在切片芋头中脂质代谢与褐变发展之间存在潜在的联系。
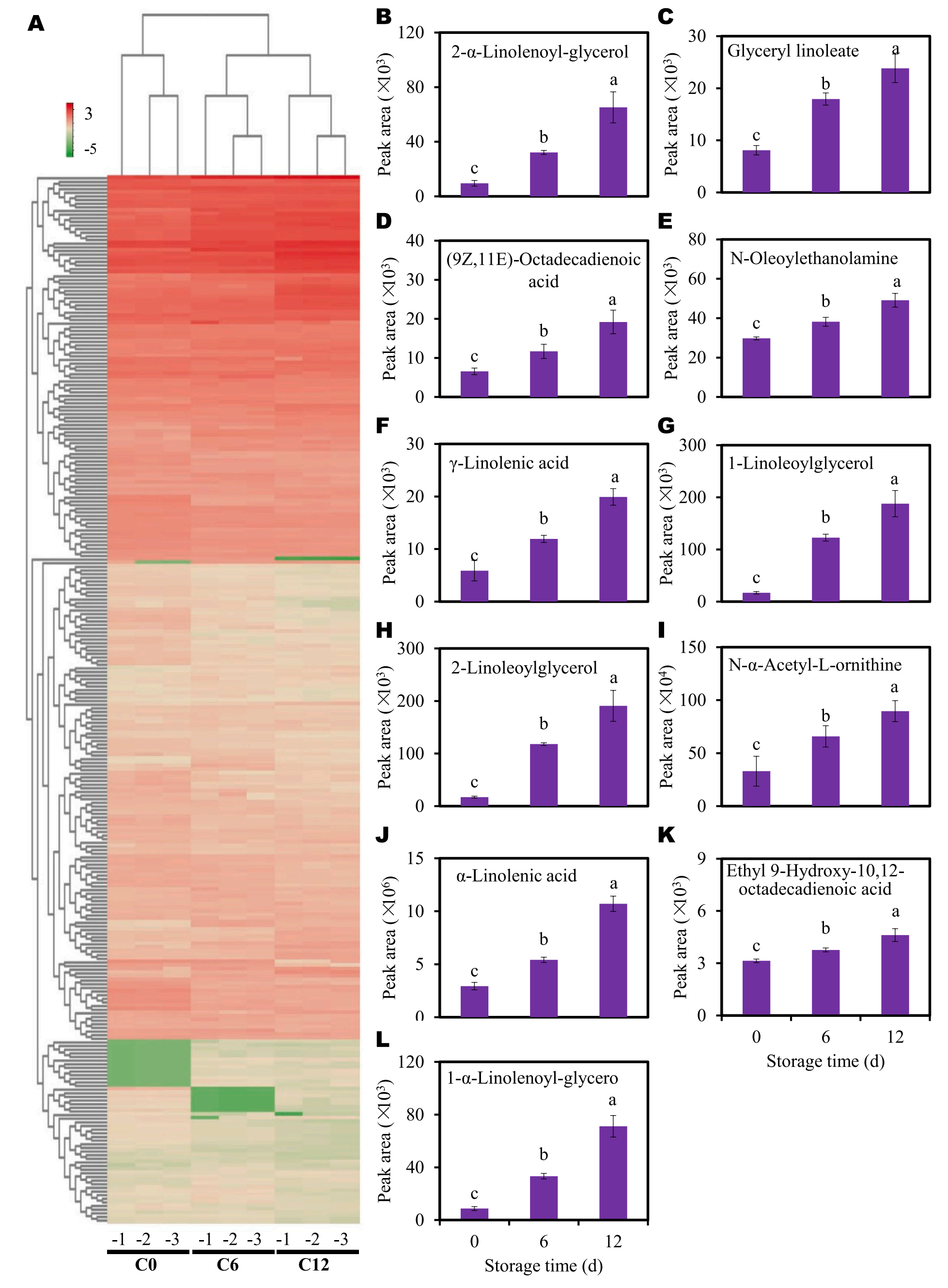
图3-切片芋头中差异代谢物(DAMs)的积累模式
4.转录组分析——冷藏芋头切片褐变过程的转录组学分析
为了更好地了解基因表达的变化,该研究进行比较转录组分析。结果显示许多基因在芋褐变过程中表现出不同的表达谱。具体来说,在C0和C6的比较中,鉴定出3103个表达上调的基因和1685个表达下调的基因。同样,在C6和C12组的比较中,发现3021个DEGs上调,2350个DEGs下调。此外,与C0组相比,C12组有5108个基因的表达量更高(图4A)。在褐变过程中发现了更多的上调基因,表明芋头的基因表达发生显著的变化。有趣的是,当比较C0与C6、C6与C12、C0与C12之间的DEGs时,发现1396个DEGs重叠(图4B)。
根据褐变过程中的表达模式,将DEGs分为6个簇。每个聚类(从1到6)分别由1607、1293、533、1741、634和2470个度组成(图4C)。KEGG富集分析显示,簇1中有四个途径的DEGs显著富集:α-亚麻酸代谢、生物素代谢、植物-病原体相互作用以及淀粉和蔗糖代谢(图4D)。此外,集群2中的DEGs在α-亚麻酸代谢、植物-病原体相互作用以及倍半萜和三萜生物合成途径中富集(图4E)。
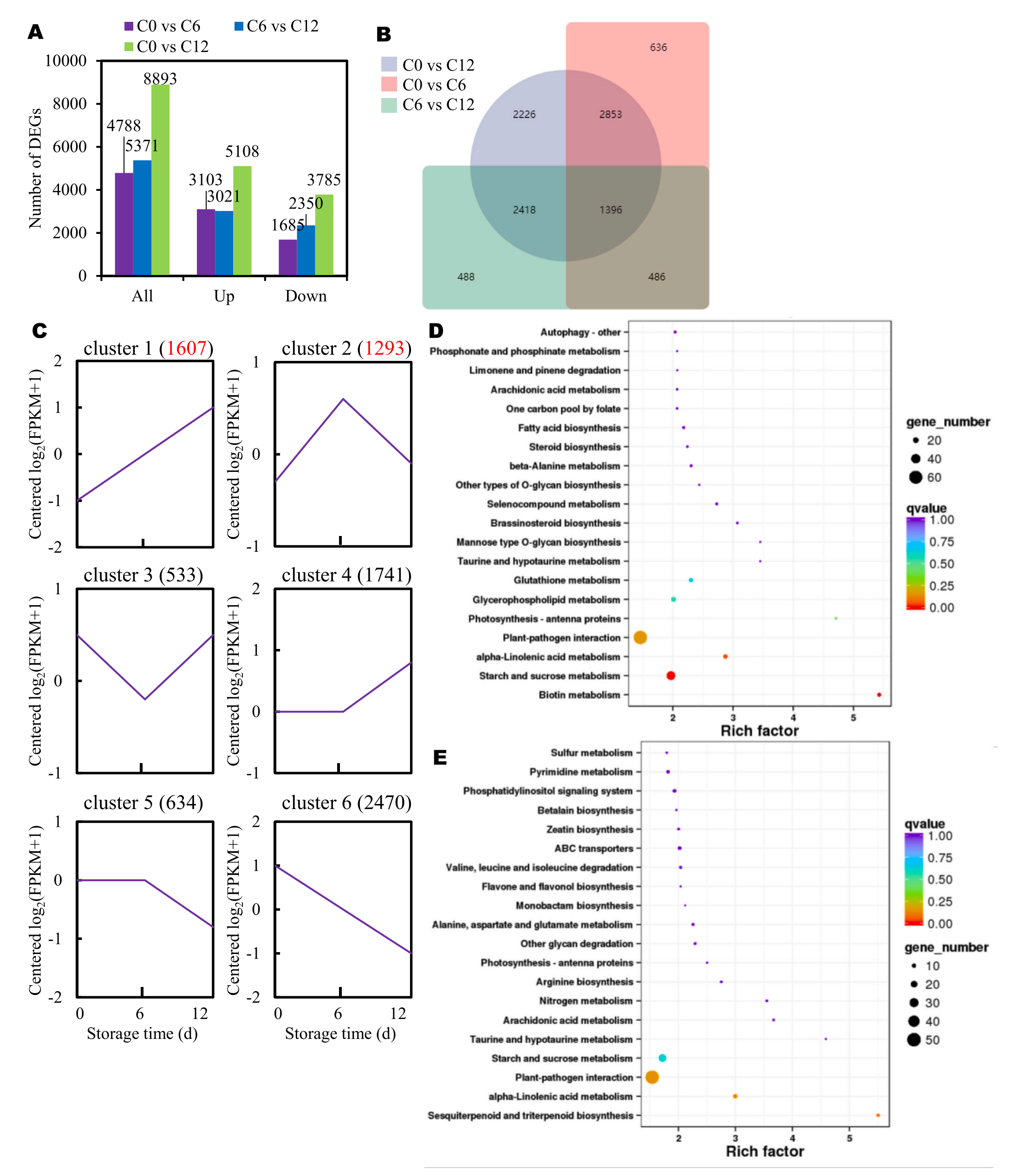
图4-切片芋头褐变过程中的转录组学分析
5.联合分析——基因共表达网络构建
为了进一步研究脂质代谢与冷藏芋头切片褐变之间的关系,研究采用WGCNA分析进行研究。如图5A所示,鉴定出两个与切片芋头在冷藏期间褐变发育显著相关的模块。蓝色模块中基因与褐变BI、a*、b*、△E 4项指标呈正相关,而绿松石模块中基因与褐变指标呈负相关(图5B)。此外,随着褐变的发展,蓝色模块中基因的表达量逐渐增加,而绿松石模块中基因的表达量逐渐减少(图5C)。此外,蓝色和绿松石模块中每个基因与褐变的相关性都很高(图5D),说明这两个模块的基因与冷藏芋头片的褐变发育高度相关。
切片芋头的褐变与蓝色模块中的基因正相关,对该模块中的基因进行了KEGG富集分析。在top20个富集的KEGG通路中,泛素介导的蛋白水解、α-亚麻酸代谢和谷胱甘肽代谢是富集最显著的通路。尤其是α-亚麻酸代谢途径和甘油脂代谢途径中分别富集了14个和18个基因(图5E)。这些结果为脂质代谢参与切片芋头褐变的事实提供了进一步的线索。

图5-WGCNA对模块与性状的相关性分析
6.基因验证——参与亚麻酸代谢的DEGs的表达模式
为了验证RNA-seq数据的质量,研究检测了亚麻酸代谢途径中DEGs的表达模式。在切片芋头褐变过程中,该途径中的DEGs差异表达(图6A)。在这些基因中,4个基因(taro_028466、029177、001379和new gene_1582)的表达量在第6天较0天下降(图6A)。10个基因(taro_026230、017933、007794、026197、032526、040173、011856、050352以及new gene_27424和3563)的表达量在第6天出现了增加,随后又出现了下降。然而,这些基因在第12天的表达水平仍然高于第0天(图6A)。其余15个基因在褐变过程中表现出稳定的表达增加。这些结果再次证实了亚麻酸代谢参与了芋头褐变过程。
作者选择了5个DEGs进行qRT-PCR分析(图6B-F)。随着贮藏时间的延长或褐变程度的恶化,这5个基因的表达量均呈上升趋势。此外,通过RNA-seq和qRT-PCR分析确定的这些基因的总体表达模式高度一致(图6B-F),证实了转录组学数据的可靠性。
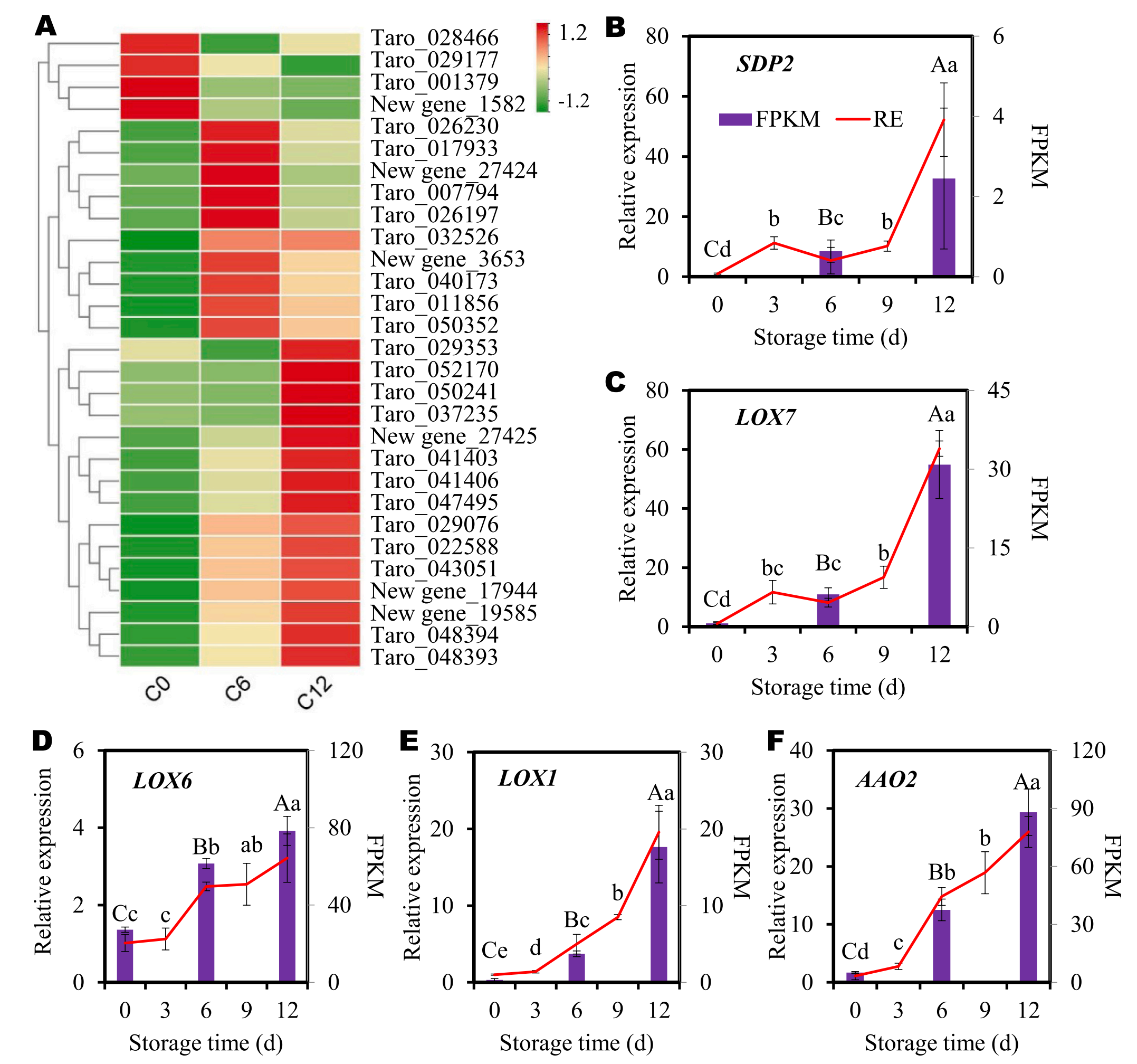
图6-亚麻酸代谢途径中DEGs的表达模式
7.基因验证——在褐变过程中,膜脂过氧化作用加剧
上述结果表明,膜脂代谢可能在芋头褐变过程中起一定作用。为了进一步研究,评估了参与膜脂过氧化的关键DEGs的表达谱,如脂肪酶(LIP)和脂氧合酶(LOX)。三个差异表达的LIP基因在褐变过程中表达持续增加。LOX基因,除了8个LOX基因在褐变过程中逐渐增加,其他基因在褐变过程中表达先增加后下降 (图7a)。这些结果表明,新鲜切割操作(如剥皮和切割)激活了芋头的脂质过氧化过程。
丙二醛通常被认为是植物脂质过氧化的生物标志物。为了监测切片芋头褐变过程中膜脂过氧化情况,测定了LOX活性和MDA含量。随着时间的推移,LOX活性和MDA含量随着褐变继续进行逐渐升高 (图7B, C)。这些结果表明,在褐变过程中,膜脂过氧化加剧。此外,相关分析显示,LOX活性、MDA含量和其他褐变指标之间存在很强的相关性 (图7D)。这些结果进一步支持了膜脂过氧化和/或代谢参与芋头褐变。
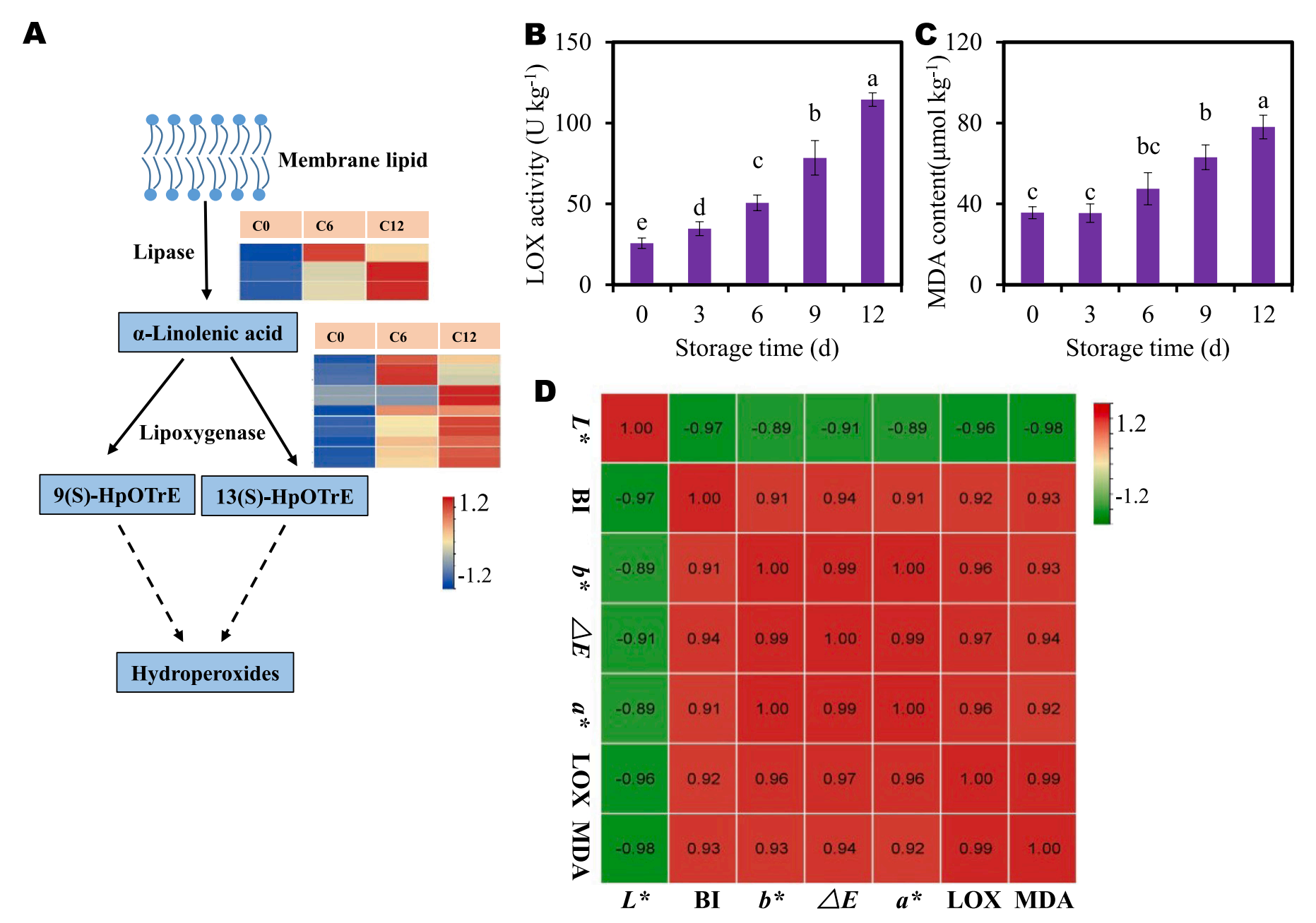
图7-膜脂过氧化的评价
研究总结
切面褐变的发生是鲜切行业中切片芋头生产和商业化的一个重要障碍。代谢组学分析显示,在芋头褐变过程中,亚麻酸及其衍生物以及氢过氧化物的含量增加,表明发生了膜脂降解。RNA-seq分析显示,参与褐变过程的DEGs富集于α-亚麻酸代谢途径。WGCNA分析得到了两个与芋头褐变密切相关的模块,其中蓝色模块的基因与BI呈正相关,强化了膜脂代谢与芋头褐变之间的联系。随着芋头褐变的进行,LOX基因的表达和蛋白活性以及MDA的水平增加,表明膜脂过氧化促进了切片芋头褐变。总之,该研究结果强调了膜脂代谢在切片芋头褐变中的重要性。这项研究通过转录组和植物广靶代谢组分析全面研究了芋头褐变的机制。




 京公网安备 11011302003368号
京公网安备 11011302003368号