

 文章标题:Chromosome-level genome assembly of Prunella vulgaris L. provides insights into pentacyclic triterpenoid biosynthesis
文章标题:Chromosome-level genome assembly of Prunella vulgaris L. provides insights into pentacyclic triterpenoid biosynthesis
期刊名称:The Plant Journal
合作单位:中国医学科学院/北京协和医学院药
研究物种:夏枯草
百迈客提供:基因组测序和部分数据分析服务
2024年1月16日,The Plant Journal上线了一篇关于夏枯草基因组的研究论文,“Chromosome-level genome assembly of Prunella vulgaris L. provides insights into pentacyclic triterpenoid biosynthesis”,报道了组装到染色体水平的夏枯草基因组,为进一步阐明五环三萜的生物合成机制奠定了基础。
摘 要
夏枯草(Prunella vulgaris)是最畅销、应用最广泛的中草药之一。据中国药典记载,它是一种清洁和保护肝脏的良药,几百年来一直被许多凉茶用作配方的主要成分。它也是欧洲和其他亚洲国家的一种传统民间药物。五环三萜是夏枯草产生的一类重要的生物活性化合物,而它们的生物合成机制仍有待阐明。本研究使用Illumina,ONT和Hi-C技术相结合的方法,获得了夏枯草的染色体水平参考基因组。其大小为671.95Mb,scaffold N50为49.10Mb,完整的BUSCO达到了98.45%。约98.31%的序列被锚定到14条假染色体上。比较基因组分析揭示了夏枯草中最近的全基因组复制(WGD)事件。全基因组分析鉴定出35932个蛋白编码基因(PCGs),其中59个编码了参与生物合成2,3-氧化鲨烯的酶。此外,还鉴定了10个PvOSC、358个PvCYP和177个PvUGT基因,其中5个PvOSCs、25个PvCYPs和9个PvUGTs被预测参与了五环三萜的生物合成。PvOSC2、PvOSC4和PvOSC6重组蛋白的生化活性测定表明,它们分别是混合香树脂醇合酶(MAS)、羽扇豆醇合酶(LUS)和β-香树脂醇合酶(BAS)。这些结果为进一步阐明夏枯草中五环三萜的生物合成机制提供了坚实的基础。
研究背景
Prunella vulgaris(图1a,b)是一种唇形科的多年生草本植物。它在中文里被命名为“夏枯草”,表明它通常在夏天枯萎。由于其在伤口愈合上的用途,P. vulgaris也被称为“self-heal”。在《中国药典》中,夏枯草的刺被记载为药用部位,并被推荐为清洁和保护肝脏的良药,特别是用于治疗肝火症状,如眼睛夜间疼痛、头晕、喉咙痛、退热。由于夏枯草具有抗微生物、抗炎症、免疫调节、抗氧化、抗肿瘤、抗高血压等多种生物活性,因此它在许多其他亚洲国家和一些欧洲国家也被用作传统民间药物。此外,在许多凉茶配方中,夏枯草也是主要成分之一。在中国,特别是在中国南方,当地人相信含有夏枯草的凉茶具有降肝火、保护肝脏免受癌症和炎症、保护呼吸系统免受喉咙痛的特性。到目前为止,夏枯草已成为中国最畅销和广泛使用的草药之一。
五环三萜及其皂苷是夏枯草中一类重要的生物活性成分,与夏枯草的抗癌和抗炎特性有关。三萜类化合物是植物中最大的次生代谢产物类别之一。根据骨架可分为无环三萜、双环三萜、三环三萜、四环三萜、五环三萜、六环三萜等亚类。三萜的生物合成从MVA途径的乙酰辅酶A和MEP途径的丙酮酸与甘油醛-3-磷酸开始。MVA途径主要作用于细胞质,在细菌、古菌与真核生物中都是保守的;而MEP途径作用于质体,主要存在于真细菌和植物中。这两种途径都导致通用C5异戊二烯单元的生物合成。它们会聚在异戊烯二磷酸(IPP)和烯丙基二磷酸(DMAPP)上。经法尼基二磷酸合酶(FPPS)、角鲨烯合成酶(SQS)和角鲨烯环氧化酶(SQE)的连续催化,IPP和DMAPP转化为三萜和甾体生物合成的关键中间体2,3-氧化角鲨烯。随后,氧化角鲨烯环化酶(OSCs)通过椅-船-椅(C-B-C)构象催化中间体环化,生成达原甾醇阳离子,用于类固醇和大多数四环三萜的生物合成,或通过椅-椅-椅(C-C-C)构象生成达玛烯基阳离子,用于生成五环三萜骨架。在植物中,循环产物的进一步修饰包括主要在细胞色素P450单加氧酶(CYPs)催化下的氧化和主要在尿苷二磷酸-糖基转移酶(UGTs)催化下的糖基化。
OSCs是由一个基因家族编码的一组环化酶,可催化2,3-氧化角鲨烯环化成结构多样的甾醇和三萜骨架。在植物甾醇(如羊毛甾醇和环阿屯醇)的生物合成过程中,OSCs通过CBC构象催化2,3-氧化角鲨烯的环化。参与这些过程的OSCs分别被称为羊毛甾醇合酶(LAS)和环阿屯醇合酶(CAS)。在五环三萜的生物合成过程中,OSCs通过CCC构象催化2,3-氧化角鲨烯的环化,根据其产物可分为混合-香树脂醇合酶(MAS)、羽扇豆醇合酶(LUS)、β-香树脂醇合酶(BAS)和DS。
CYP超家族是植物中最大的基因超家族之一。根据CYP系统发育树的分支,植物CYP蛋白可分为11个氏族和127个家族。有七个氏族是单家族,包括CYP51、CYP74、CYP97、CYP710、CYP711、CYP727、CYP746,而其余的CYP71、CYP72、CYP85、CYP86氏族为多家族。在生成骨架后,在五环三萜的C-2、C-3、C-16、C-19、C-22、C-23、C-24、C-28等不同位点进行氧化和糖基化修饰,生成大量的三萜。氧化修饰主要涉及CYP716家族成员。此外,CYP51、CYP71、CYP72、CYP85、CYP87、CYP88和CYP93家族的一些成员也可能参与。
以尿苷二磷酸糖作为糖基供体的可能性命名的UGTs,在已发现的106个糖基转移酶家族中属于第1家族。根据UGTs的3D结构状态,将其分为GT-A、GT-B、GT-C和GT-D四种类型。植物UGTs属于GT-B,它在开花植物中可进一步分为18个类群(A-R)和一个外类群。五环三萜的糖基化通常发生在C-3、C-23和C-28位点,主要涉及UGT73家族成员和UGT71、UGT74、UGT85、UGT91和UGT94家族的部分成员。虽然C-19羟基化的五环三萜类化合物,如苦委陵菜酸(TA)、坡模酸(PA)及其皂苷广泛存在于植物中,但迄今为止,负责C-19羟基化的氧化酶尚未被发现。
虽然夏枯草具有很高的药用价值,但与农作物、模式植物以及丹参、人参和罂粟等多种药用植物相比,对其研究较少。关于其生物活性成分及其生物合成机制的报道很少。为了促进对夏枯草的研究,我们采用Illumina、ONT和Hi-C技术相结合的方法对其基因组进行了测序,并将序列组装到染色体水平,然后进行比较分析。参与五环三萜生物合成的基因在全基因组范围内被鉴定出来,并揭示了三种PvOSCs的功能。这些结果为进一步分析提供了夏枯草属的第一个染色体水平的基因组。
研究结果
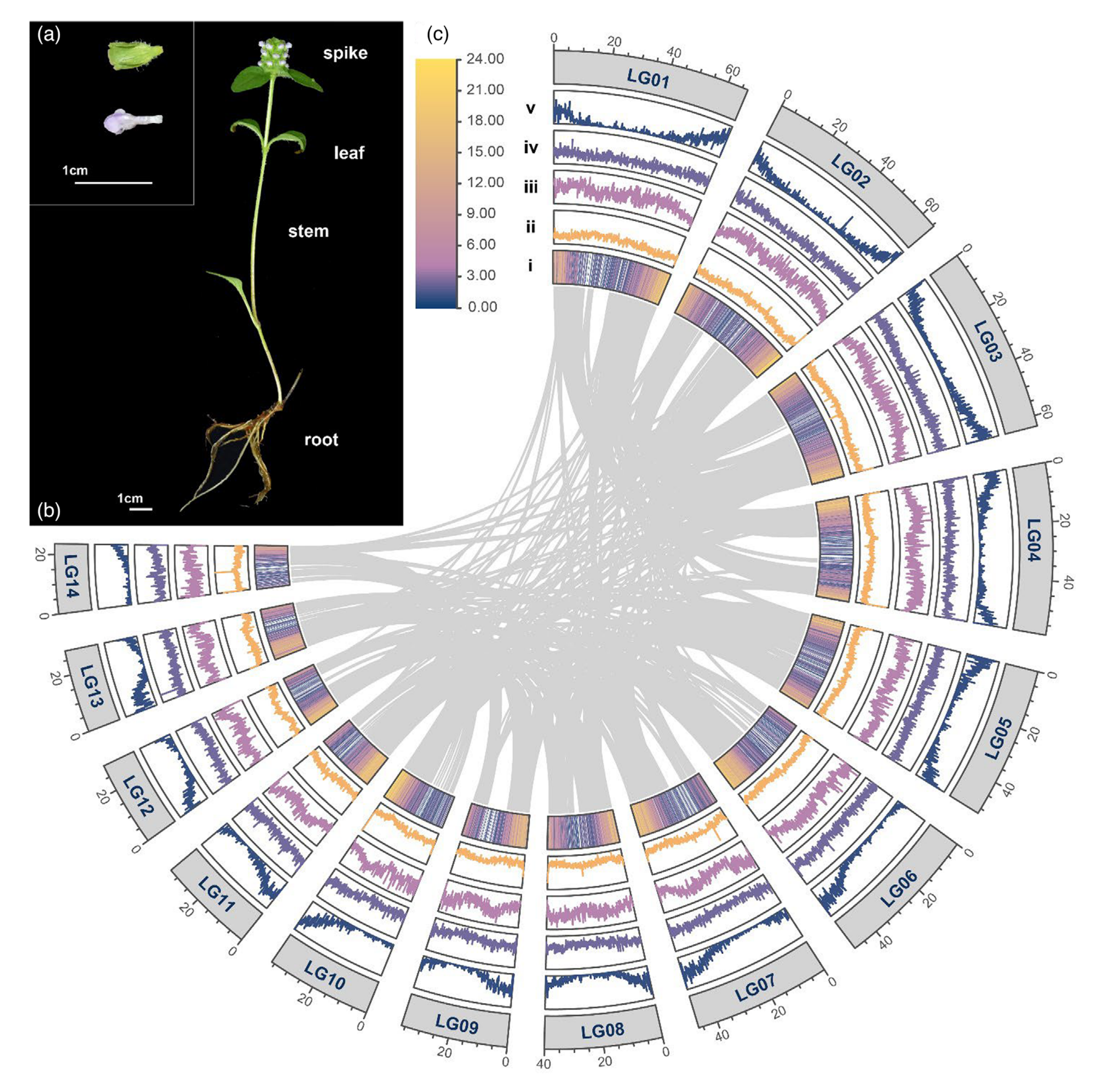
图1-夏枯草的形态、基因组和基因组内共线性概览
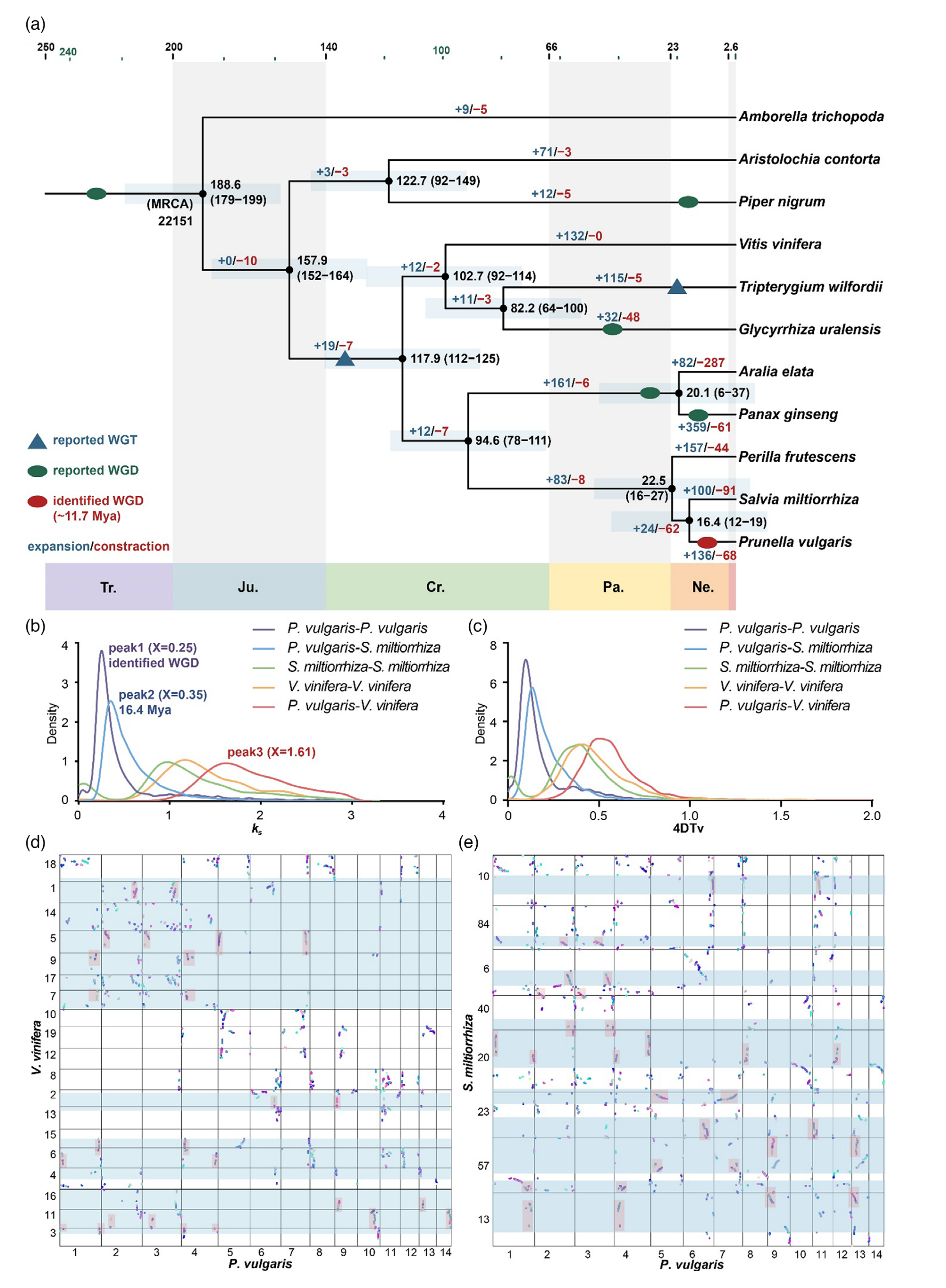
图2-比较基因组学分析
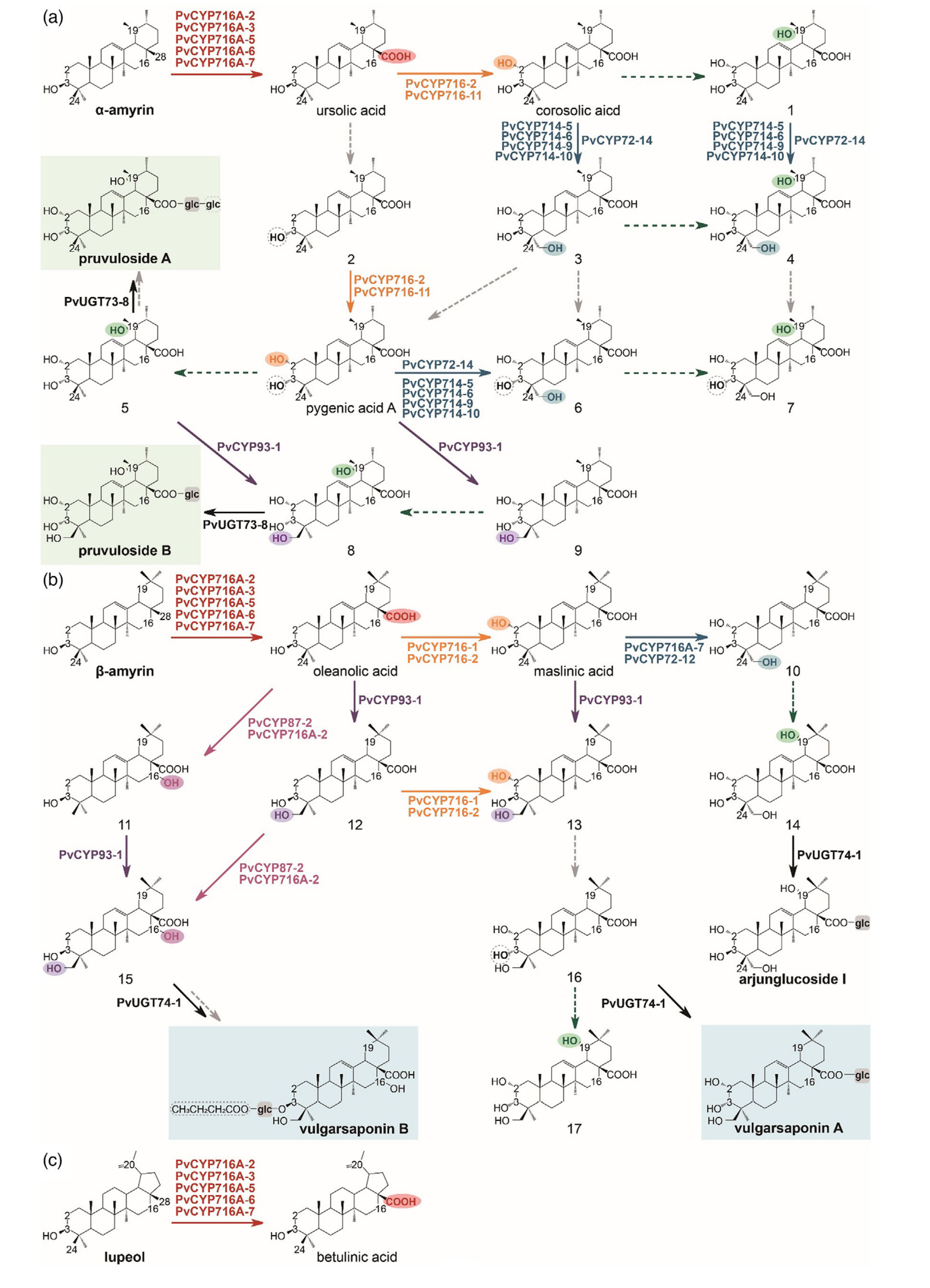
图3-夏枯草中α-/β-香树脂烷型和羽扇烷型五环三萜的生物合成途径
参考文献:Zhang, S., Meng, F., Pan, X., Qiu, X., Li, C. and Lu, S. (2024), Chromosome-level genome assembly of Prunella vulgaris L. provides insights into pentacyclic triterpenoid biosynthesis. Plant Journal. https://doi.org/10.1111/tpj.16629
文章来源于植物代谢研究




 京公网安备 11011302003368号
京公网安备 11011302003368号