

关于植物应对盐碱胁迫的内部调控机制的研究一直以来是生物研究的热点内容,对胁迫的信号传递和应答过程的深入了解将有助于提高作物的逆境适应能力,21年百迈客转录调控合作案例300+,影响因子1200+,其中植物的胁迫研究独占鳌头,今天给大家分享一篇镉胁迫研究方向的好文解析。

英文标题:CadmiumbindingduringleafsenescenceinFestucaarundinacea:Promotionphytoextractionefficiencybyharvestingdeadleaves
发表期刊:CHEMOSPHERE
影响因子:7.086
全文链接:https://international.biocloud.net/zh/article/detail/34902388
研究背景
枯叶提取法是一种新型的高羊茅镉植物修复策略,为镉污染土壤的植物修复和清洁食品生产提供了可行性。枯叶中Cd含量最高是叶片衰老过程中Cd积累的结果。然而,叶片衰老过程中Cd积累的机理尚不清楚,这限制了该技术的提取效率。本研究通过RNA-seq组学研究技术对高羊茅叶片Cd积累的内部机制。
研究方法
根据叶片在美洲虎羊茅茎中的位置进行取样和分离。L1为顶部初生叶,L2和L3为上部2、3个成熟叶片,L4是衰老开始时的叶片;L5是衰老的黄色叶片。
分别用75μMCd和不加75μMCd处理高羊茅幼叶(L1)和衰老叶(L4)7d(L1-con、L4-con、L1-Cd、L4-Cd),并进行3次生物重复,提取总RNA并进行转录组测序。(测序工作由北京百迈客公司提供技术支持)。
研究结果
一、不同叶片中的镉、游离氨基酸、可溶性糖和金属硫蛋白测定
不同叶片Cd积累量显著不同。叶片Cd浓度随叶片数量的增加从上到下显著增加,L2、L3、L4和L5的Cd浓度比幼叶(L1)增加了2.1、2.3、2.5和5.1倍。幼叶(L1)积累Cd最少,而衰老叶(L5)积累Cd最多。Cd处理显著提高了各叶片游离氨基酸含量,但没有改变其随叶片衰老逐渐下降的趋势,幼叶中游离氨基酸含量(L1)最高,然后随着叶片衰老逐渐降低(图1B)。衰老叶(L5)中游离氨基酸含量最低。成熟叶片中可溶性糖含量(L2)最高,Cd处理显著提高了L2和L4的可溶性糖含量,然后随着叶片衰老逐渐降低。衰老叶(L5)和幼叶(L1)的可溶性糖含量显著低于绿色成熟叶。镉胁迫下,与对照组相比,Cd处理显著提高了所有叶片中的金属硫蛋白含量,不同叶片金属硫蛋白含量无显著变化。
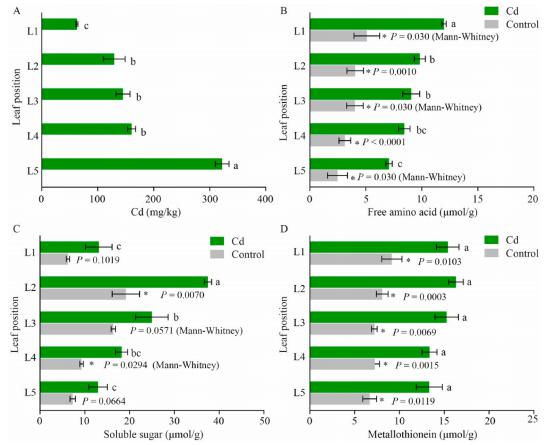
Cd胁迫对高羊茅不同叶片Cd(A)、游离氨基酸(B)、可溶性糖(C)和金属硫蛋白(D)含量的影响
二、镉处理对不同叶片间螯合物的影响
PC、GSH和NPT参与了超积累剂的Cd解毒机。Cd胁迫和对照组的植物螯合素(PCs)均随叶片老化显著增加。Cd处理显著提高了各叶片PC含量,非蛋白硫醇(NPTs)含量也随着叶片衰老而显著增加,但随叶片衰老的增加趋势没有变化。Cd处理显著增加了衰老叶片(L4和L5)的谷胱甘肽(GSH)含量,但没有提高幼苗和成熟苗的GSH含量。由此可以推断,GSH、PC和NPT的增加及其与Cd的结合可能促进了衰老叶片中Cd的积累。Cd处理显著增加了幼叶和成熟叶的氧化谷胱甘肽(GSSH)含量(L1和L2),但降低了衰老叶(L5)的GSSH含量。由于氧化后的谷胱甘肽(GSSH)可被还原为还原型谷胱甘肽(GSH),还原性谷胱甘肽(GSH)可以通过GST进一步催化GSH-Cd结合,GSSH向GSH的转化与Cd在衰老叶片中的积累是一致的。
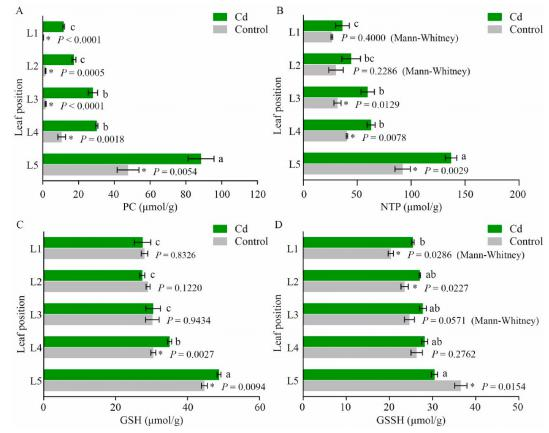
Cd胁迫对高羊茅不同叶片螯合素(PCs)、非蛋白硫醇(NPTs)、还原型谷胱甘肽(GSH)、氧化型谷胱甘肽(GSSH)含量的影响
三、差异基因的鉴定和GO分析
为了更好地了解Cd在叶片衰老过程中的积累机制,利用RNA-seq研究了Cd处理前后幼叶(L1)和衰老叶(L4)的转录组谱。对L1-ConvsL4-Con、L1-CdvsL4-Cd、L1-convsL1-Cd和L4-convsL4-Cd的比较组根据转录水平至少有两倍差异进行筛选,分别发现的差异基因数目为2286、4270、1808、1929。在这些差异基因中,与Cd胁迫相比,L1和L4中有157个基因表达上调,255个基因表达下调。四个比较组的共有24个上调基因和20个下调基因。采用qRT-PCR方法,从RNA-seq数据的差异基因中随机选取12个基因的表达水平进行分析。这12个基因在qRT-PCR分析结果和RNA-seq数据中均显示出相同的表达模式。
为了了解差异基因的功能,作者进行了GO富集分析。四组的差异基因在节律过程、信号传导和代谢过程等生物过程中都有20个丰富的分类。值得注意的是,与对照组相比,脱毒组的差异基因在幼叶和衰老叶中都有富集(L1-ConvsL1-Cd和L4-ConvsL4-Cd),但在幼叶和衰老叶的对照组和处理组中没有明显富集(L1-CdvsL4-Cd和L1-ConvsL4-Con)。
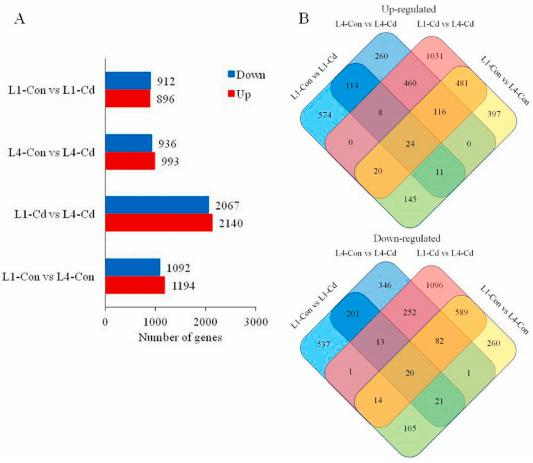
镉胁迫下幼叶和衰老叶片基因表达谱的变化
四、KEGG通路分析
为了进一步了解Cd处理下高羊茅叶片中Cd积累的机制,利用KEGG对叶片中的差异基因进行富集分析。在幼叶中,主要富集的途径是昼夜节律-植物、苯丙氨酸代谢、植物激素信号转导、苯丙素生物合成、淀粉和蔗糖代谢,而在衰老叶片中,差异基因主要富集于类胡萝卜素的生物合成、昼夜节律与植物的关系、谷胱甘肽的代谢、植物与病原菌的相互作用以及苯丙素的生物合成等代谢途径。苯丙素类生物合成和昼夜节律型植物的基因在幼叶和衰老叶中都表现丰富。值得注意的是,属于谷胱甘肽代谢途径的基因仅在衰老叶片中表现出富集。

差异基因的KEGG途径富集分析
五、衰老叶片中谷胱甘肽代谢途径与Cd结合
通过比较分析幼叶和衰老叶片对Cd胁迫的响应,有助于探索衰老叶片中Cd积累的关键调控基因,作者进一步比较了对照组组和Cd处理组在“谷胱甘肽代谢”途径中的差异基因表达。谷胱甘肽-S-转移酶(GSTs)是催化GSH-Cd偶联的酶,植物GSTs可以进行GSH依赖的反应来进行Cd解毒。在该转录组中,共鉴定出了26个基因编码GST。其中,19个上调基因在Cd胁迫下的差异倍数高于对照组。结果表明,GST基因的上调和S-谷胱甘肽的结合可能参与了衰老叶片中Cd的积累。
MT(金属硫蛋白)是一组金属结合蛋白,在金属分配和稳态中起着关键作用。1个上调基因(c129583.Graph_c0)和一个下调基因(c120767.graph_c0)编码MT。Cd胁迫使上调基因的差异倍数比对照增加了23.8%,并完全抑制了表达下调的基因。结合Cd胁迫下MT含量的增加,作者的研究结果表明,MT-Cd结合可能在幼叶(L1)和衰老叶片(L4)的Cd解毒过程中发挥作用。半胱氨酸蛋白酶(CPs)是叶片衰老过程中负责蛋白水解活性的最丰富的酶。在该转录组中鉴定出17个编码CP的基因。在对照组中,只有4个基因的表达有显著差异。Cd胁迫显著促进CP表达,有10个上调基因在Cd处理组的差异倍数高于处理组,Cd处理组的6个基因下调倍数低于对照组。然而,PC的蛋白水解活性是否与衰老叶片中Cd的积累有关尚不清楚。
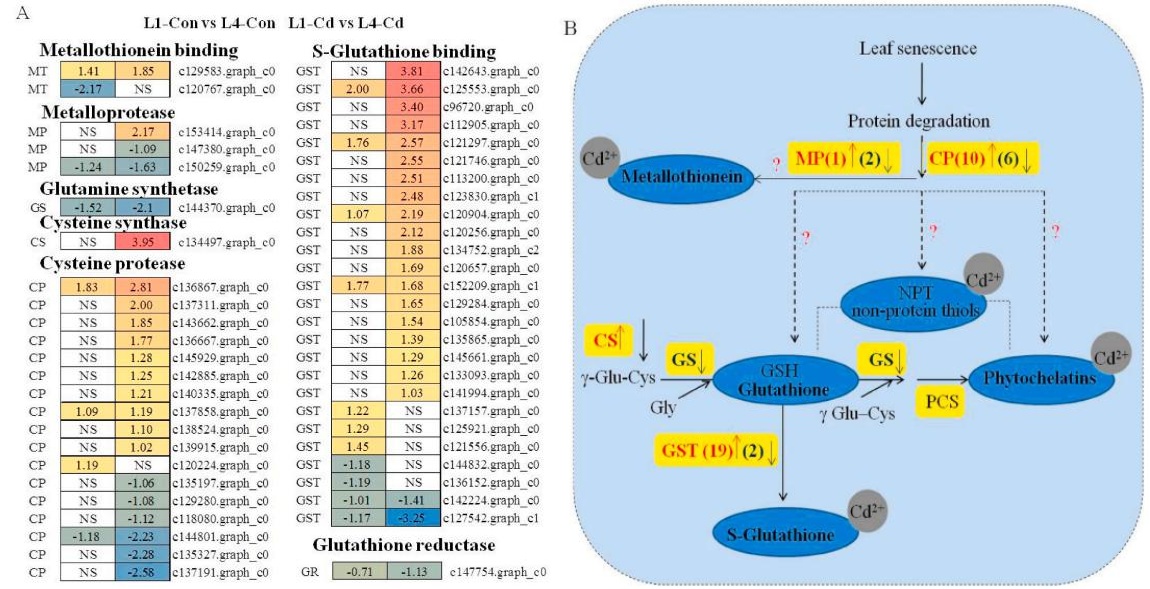
与谷胱甘肽代谢和Cd结合相关的基因在幼叶(L1)和衰老叶(L4)与Cd胁迫或不胁迫下的转录变化
总结
从枯叶中提取Cd的策略取决于叶片衰老过程中Cd的积累机制。研究叶片衰老过程中Cd在枯叶中优先积累的机理至关重要。本研究通过转录组和化学分析表明,叶片衰老过程中Cd的积累与PC、GSH和NPT结合Cd有关。Cd结合PC、GSH、NPT的增加可能来自于蛋白质降解的途径。作者的研究结果为揭示叶片衰老过程中Cd积累的调控机制提供了依据。在高羊茅蛋白质降解过程中,通过促进PC、GSH、NPT与Cd的结合,可进一步提高高羊茅枯叶提取效率。




 京公网安备 11011302003368号
京公网安备 11011302003368号