


中文题目:影响耐冻性的因素:越冬常绿植物田间和人工低温驯化之间的转录组学比较研究
发表期刊:thePlantJournal
发表日期:2020.06
影响因子:5.901
研究背景
抗冻性不仅决定着植物的地理分布,而且影响着作物、蔬菜和水果的产量,已成为温带和北温带地区人们关注的焦点。植物通过一个称为“低温驯化”的过程来增加它们的抗冻性,而通过“脱驯化”失去抗冻性。低温驯化是一个多基因调控的过程,伴随着植物的生理调节。多年生木本植物的低温驯化通常包括两个阶段,分别由短光周期和低温诱导。尽管多年生木本植物对光周期和低温的表型反应已经被了解得很清楚,但在分子水平上知之甚少。
在这篇文章中,研究者选用多年生木本植物杜鹃花属的常绿品种‘ElsieLee’进行低温驯化,其叶子在冬天变为红色,春天又变成绿色,这为花青素在低温驯化中的重要作用提供了机会。本研究结果可全面认识野外和人工低温驯化的差异,为了解多年生木本植物越冬常绿叶片的低温驯化奠定基础。
研究方法
选3年生的“ElsieLee”杜鹃,用于田间或人工低温驯化试验,共3组试验:ExperimentI田间低温驯化(F-CA),ExperimentII人工低温驯化(A-CA),ExperimentIII人工用紫外光B进行低温驯化(A-CA+UVB)。每组试验36盆,分为3个生物学重复,每个生物学重复12盆,采集当年生长的完全展开的叶子进行实验,叶片在抽样日期收集后立即在液氮中冷冻,然后-80℃保存。
研究结果
1、叶片耐冻性及花青素的积累
LT50为引起50%损伤的温度,在试验I中,10月26日(该日期下的样品视为未驯化的样品)平均温度约为22℃,光照11.2h,此时LT50为-4.3℃。试验II和III在人工低温驯化前处在营养生长阶段,温度为25℃,光照16h,LT50为-4.2℃。
F-CA(10月26日-1月20日)期间,LFT随温度降低和光照减少而增强,1月20日,叶片组织获得整个F-CA期最强的LFT(LT50为-18.5℃)。A-CA(4℃)或UVB(4℃+紫外光B)驯化56天后,LFT提高至-10℃。F-CA中,叶片花青素含量逐渐增加,但在A-CA中没有明显变化,而在UVB中,处理14天后,花青素开始增加,处理56天后,花青素的浓度约为1月20日F-CA处理时的一半。结合A-CA和UVB的LFT数据,花青素本身是否增强了LFT尚不清楚。
2、转录组测序、主要成分分析,以及在田间和人工低温驯化过程中差异表达基因的鉴定
根据LFT和花青素的生理变化,选择5个样本进行转录组分析,每个样本三个生物学重复(图1a)。
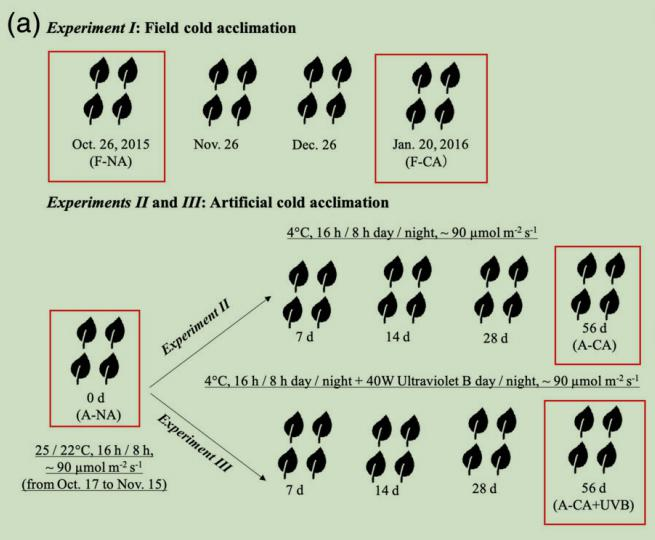
图1a田间和人工驯化的样品采集
转录组共鉴定出94,144个unigenes,N50为1800。所有unigenes被用来进行主成分分析(图2a),结果显示(i)每个样本生物学重复功能上相似;(ii)主成分1(PC1)能将CA与非驯化分离,主成分2(PC2)能够将F-CA与A-CA和A-CA+UVB分开;(iii)F-NA与A-NA的距离表明叶片在早秋与营养生长时的状态是不同的。

图2(a)RNA-seq鉴定的所有unigenes的主成分分析,(b)DEGs差异表达基因的韦恩图,(c)基于DEGs的KEGG通路的韦恩图
三组试验F-NAvs.F-CA(I)、A-NAvs.A-CA(II)及A-NAvs.A-CA+UVB(III)分别注释了3023、1235和2620个差异表达基因,其中438个为共同差异表达基因,另有2229、122和1259个上调或下调基因分别为试验I、II和III所特有的(图2b)。
试验I、II和III富集的KEGG通路分别为23、26和31个(图2c)。其中8个为试验I特有的,包括4个光合作用相关通路(KO00195、KO00196、KO00906和KO04146)及两个脂肪酸代谢通路(KO00591和KO00592)。三组试验共有的通路为11个,包括5个碳水化合物通路(KO00040、KO00053、KO00500、KO00630和KO01200)、3个次级代谢通路(KO00904、KO00940和KO00941)、1个氨基酸代谢通路(KO00410)、1个脂质代谢通路(KO00071)和1个昼夜节律通路(KO04712)(图2c)。
3、在F-CA中的特异性富集的KEGG通路
KO00906为类胡萝卜素生物合成途径,在试验I中富集(图2c),该途径导致ABA的生物合成和分解代谢(图3a)。该通路中,3个被注释为β-胡萝卜素3-羟化酶(CHY-β)表达量上调(图3b)。CHY-β将β-胡萝卜素转化为玉米黄素,玉米黄素在非光化学猝灭的热耗散中起关键作用。一个被注释为玉米黄素环氧化酶(ZEP)的差异基因表达下调。在叶黄素循环中,ZEP将玉米黄素/环氧玉米黄素转化为紫黄素。因此,在试验I中,玉米黄素的积累可能与CHY-βs上调及ZEP下调有关(图3b)。
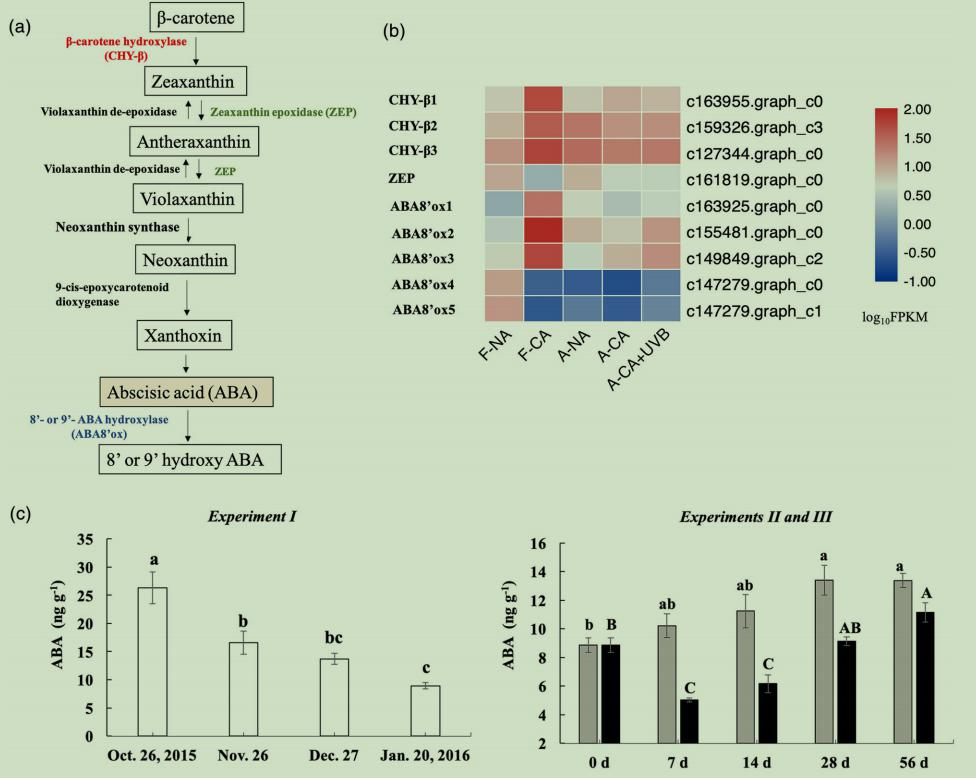
图3(a)β-胡萝卜素介导的脱落酸代谢途径,(b)β-胡萝卜素介导的脱落酸代谢途径的DEGs热图,(c)田间(试验I)和人工(试验II和III)低温驯化过程中叶片组织ABA含量的变化。
有趣的是,有5个注释为ABA8-羟化酶(ABA8’ox),其中在F-NA中表达水平较低的ABA8’ox1-3在F-CA中表达上调,而ABA8’ox4-5在F-CA中受到抑制(图3b),类胡萝卜素是ABA的前体,而ABA是ABA8’ox的底物(图3a)。ABA8’ox随内源ABA水平的增加而上调。试验I10月26日的ABA浓度是试验II和试验III0天处理的3倍左右(图3c),试验I1月20日的ABA水平下降到试验II和试验III0天处理的水平(图3c)。因此,ABA8’ox1-3可能参与了F-CA过程中ABA的分解代谢,而ABA8’ox4-5可能在非驯化条件下受高水平ABA诱导(图3c)。
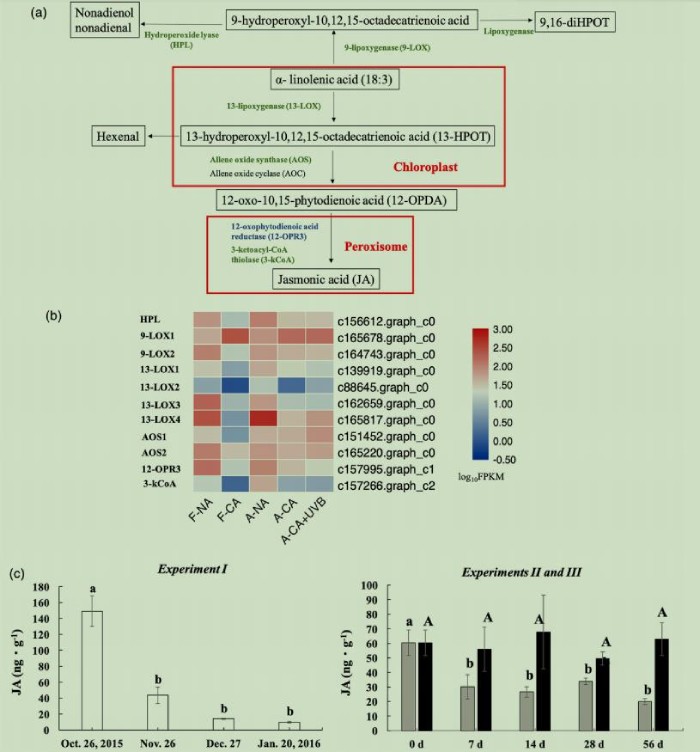
图4(a)亚麻酸代谢——茉莉酸的生物合成途径,(b)与这一途径相关DEGs热图,(c)田间(试验I)和人工(试验II和III)低温驯化过程中叶片组织JA含量的变化。
试验I中,KO00591亚油酸代谢和KO00592α-亚麻酸代谢分别富集了7个和20个DEGs。这些途径促进JA的生物合成(图4a)。5个编码脂氧合酶(LOX)的DEGs同时存在于这两个途径中且表达下调。α-亚麻酸和亚油酸都是多不饱和脂肪酸(PUFAs)和LOX的底物。α-亚麻酸代谢途径的20个DEGs中有7个是上调的。其中5个被注释为12-氧代植二烯酸还原酶(12-OPR3),转录水平很低。另一个注释为12-OPR3的基因在F-NA中高表达,而在F-CA中表达下调(图4b)。
试验I的茉莉酸含量在10月26日至1月20日期间大幅度下降了93%,试验II在0~56天期间下降了66%;而在试验III中,没有明显变化(图4c)。AtLOX2和AtCYP74A(alleneoxidesynthase,AOS)已被证明是拟南芥JA生物合成的关键酶,其中任何一种的缺失都会抑制JA的生物合成。此外,NaLOX3参与了烟草中JA的积累。本研究中,13-LOXs和AOSs在试验I和II中均下调(图4b),表明抑制亚麻酸的消耗可能是为了在应对低温时保持膜的流动性。UVB被证明可以诱导拟南芥中JA的生物合成。因此,低温抑制JA生物合成,而UVB处理可能导致整个试验III中JA含量稳定(图4c)。
KO00195光合作用、KO00196光合作用-天线蛋白和KO04146过氧化物酶体通路在试验I中特异性富集(图2c),它们与光保护相关。在KO00196光合-天线蛋白通路中,DEGs被注释为光捕获复合物,9个DEGs中有4个表达下调,这4个下调的DEGs的转录水平(即FPKM值)均高于F-NA中的其他DEGs,表明这4个基因在植物中起着关键作用,冬季叶片组织试图通过减少光收集复合物来降低对光能的吸收。
试验I中叶绿素(a+b)含量从10月26日至1月20日(图5a)降低了30%(图5a),试验II和III在0-56天处理后分别降低了23%和15%(图5b)。这表明,随着温度的降低,叶片试图通过降低叶绿素含量来降低光吸收。试验I中,叶绿素a/b在10月26日至1月20日下降了15%(图5c);试验II和III中,从0-56天分别减少8%和6%(图5d)。叶绿素a/b的变化表明试验I反应中心的降解比试验II和III更严重。代表光系统II反应中心开放度的光化学猝灭(qP)在10月26日至1月20日,降低了57%(图5e),试验II和III处理0~56天后,qP分别降低了43%和25%(图5f)。叶绿素(a+b)、叶绿素a/b和qP在试验I中下降得更多,说明光保护或光抑制作用更明显。
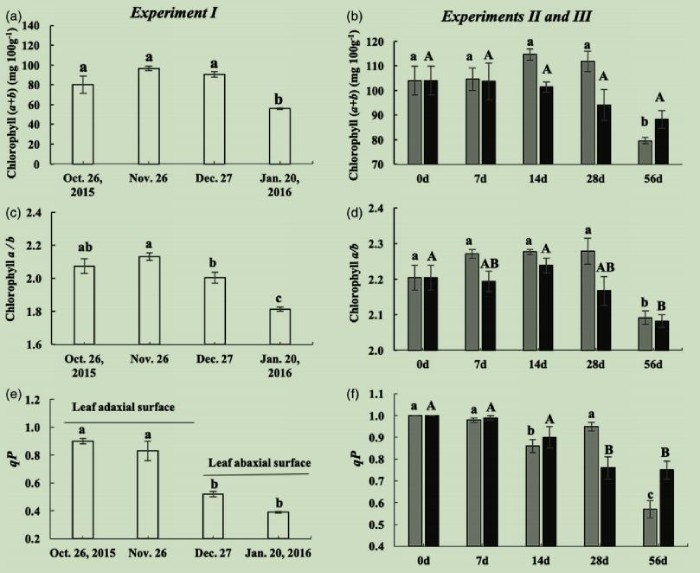
图5叶绿素(a+b)(a,b)、叶绿素a/b(c,d)和光化学猝灭(e,f)在试验I、II和III中的改变。
在KO00195光合作用通路中,DEGs被注释为负责电子传递、光系统I或II亚基以及ATP酶等活性蛋白。在KO04146过氧化物酶体通路中,8个被注释为超氧化物歧化酶和过氧化氢酶的DEGs表达上调。这两种途径中大部分DEGs的转录水平较低。
此外,除KEGG通路中富集的DEGs外,还有3个编码早期光诱导蛋白(ELIPs)的DEGs。ELIPs在试验I中被高度诱导表达,且表达上调的幅度比试验II和III中要高。ELIPs定位于类囊体膜,它们结合叶绿素a并在光保护中发挥重要作用。它们的积累可以被各种生理条件触发,包括光胁迫和低温,并与光系统II反应中心降解相关。试验I中ELIPs的表达显著上调,说明F-CA中光保护或光抑制作用更明显。
4、在F-CA和A-CA中普遍富集的KEGG通路
(1)碳水化合物的新陈代谢。属于碳水化合物代谢的5个KEGG通路在三个试验中都有富集(图2c和图6a)。
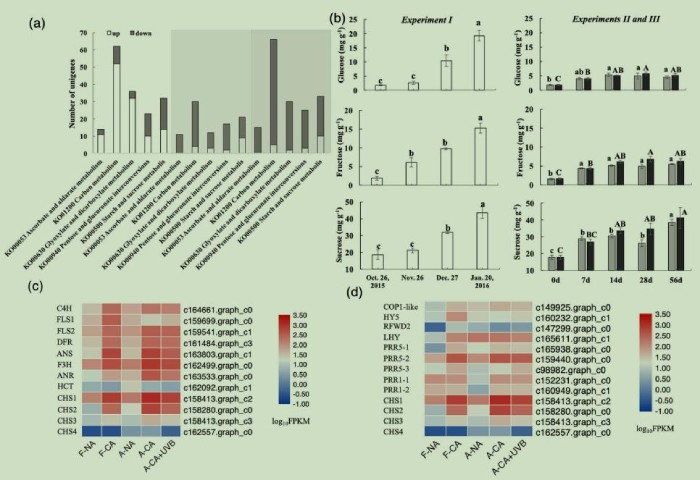
图6(a)试验I(白色背景)、II(浅灰色背景)和III(深灰色背景)中普遍富集的碳水化合物代谢通路(KEGG通路,p<0.05),(b)试验I、II和III在低温驯化期间叶片中葡萄糖、果糖和蔗糖浓度的变化,(c)KO00941类黄酮生物合成通路的DEGs热图,(d)KO04712植物昼夜节律通路的DEGs热图
KO00053抗坏血酸和aldarate代谢、KO01200碳代谢、KO00630乙醛酸和二羧酸代谢途径中,大部分DEGs在试验I中表达上调,而在试验II和试验III中表达下调(图6a)。在KO00040戊糖和葡萄糖醛酸转化途径中,试验I的23个DEGs中有10个上调,试验II的17个DEGs中只有2个上调,试验III的25个DEGs中有3个上调(图6a)。在KO00500淀粉和蔗糖代谢中,试验I的32个DEGs中有14个上调,试验II的21个DEGs中有9个上调,试验III的33个DEGs中有10个上调(图6a)。KO01200碳代谢图(未展示)显示,试验I中磷酸戊糖途径的所有DEGs和糖酵解过程中的大部分DEGs均上调,但在试验II和试验III中均下调。磷酸戊糖途径和糖酵解途径属于呼吸途径,在碳水化合物代谢中起重要作用。这些数据表明,试验I的碳水化合物代谢变化与试验II和III不同,这可以通过糖含量的变化来证明(图6b)。
试验I中,10月26日至1月20日,叶片葡萄糖和果糖浓度分别增加了10倍和7倍(图6b)。然而,试验II和III中,从0-56天处理,葡萄糖和果糖分别仅增加了~1.6倍和2.7倍(图6b)。试验I在10月26日至1月20日期间叶片蔗糖浓度增加了1.3倍,试验II和III在0~56天期间分别增加了1.2倍和1.3倍(图6b)。这些结果表明,葡萄糖和果糖在抗冻性中比蔗糖更重要。
(2)花青素生物合成。KO00941类黄酮生物合成途径在试验I、II和III中富集的DEGs分别为8、8和11个,这些DEGs也参与了花青素的生物合成。试验I中8个DEGs均上调,试验II和III中分别有1个和3个DEGs表达下调(图6c)。除FLS1、HCT和CHS4外,F-NA和A-NA中其他DEGs的FPKMs均低于低温驯化样品(即F-CA、A-CA和A-CA+UVB;图6c)。虽然花青素在A-CA中没有显著积累(图1e),但C4H、FLS2、DFR、ANS、F3H、ANR、CHS1、CHS2和CHS3的转录水平(图6c)高于A-CA+UVB或F-CA,说明低温可以诱导花青素生物合成相关基因的表达,而花青素生物合成可能是一个转录后调控的过程。
花青素在细胞质中合成,然后通过谷胱甘肽S-转移酶(GST)转移到液泡中,GST催化花青素与谷胱甘肽结合。在本研究中,KO00480谷胱甘肽代谢途径在试验I和III中富集并伴随花青素积累(图1e,2c)。在试验I和III中,分别有18和19个DEGs,其中分别有12个(都表达上调)和13个(其中8个表达上调)被注释为GST。说明在花青素合成后谷胱甘肽代谢是一个重要的途径。
(3)KO04712植物昼夜节律通路。该通路在试验I、II和III中分别富集了8、6和8个DEGs,除了试验III中的一个下调外,其余都上调。COP1-like、HY5和LHY的差异表达只存在于试验I中(图6d)。COP1在F-NA和A-NA中呈现不同的初始转录水平,并在试验I、II和III中上调(图6d)。COP1转录后影响HY5表达。HY5是碱性亮氨酸拉链(bZIP)家族的转录因子,是多个光感受器的下游,该基因在转录上受光照或低温的影响。HY5在试验I中表达量显著上调,而4℃处理后,在试验II和III中没有显著变化(图6d),表明HY5的调控可能比以前报道的更复杂。LHY是MYB相关转录因子,被归类为生物钟的晨起基因。在本研究中,LHY在F-NA和A-NA中的初始表达水平显著不同,试验I中该基因表达上调,而在试验II和III中表达差异不显著(图6d)。其他的DEGs被注释为伪响应调节器(PRR)/类双组分响应调节器或低温相关查尔酮合成酶(CHS),普遍存在于试验I、II和III中(图6d)。PRR与LHY在生物钟中被归为晨起基因,LHY的表达增强了PRR的表达。试验II中昼夜节律通路的DEGs表明,低温本身干扰了植物的昼夜节律。
结论
研究者通过3个试验来表征越冬常绿叶片低温驯化的特征,并发现低温前的光信号对于进一步增加耐冻性至关重要,因为某些信号转导通路的激活可能会导致生长停止和低温驯化。玉米黄质的积累是ABA生物合成的上游环节,可能在ABA水平中起关键作用。在本报告中,与营养生长的植物相比,ElsieLee的ABA可能在早秋通过对光信号的响应而积累,在冬季到来时下降。光信号和低温均扰乱了ElsieLee的昼夜节律,转录因子HY5和LHY均参与其中。此外,F-CA和A-CA在激素信号转导、光抑制/光保护程度、脂肪酸代谢和呼吸等方面的过程也有很大的不同,导致完成低温驯化或未完成低温驯化时的耐冻性不一致。UVB处理A-CAs表明,花青素在常绿叶片低温驯化过程中与光保护的相关性大于增加耐冻性的相关性。




 京公网安备 11011302003368号
京公网安备 11011302003368号