

明星基因SHR、SCR又发《Cell》子刊了?这到底是怎么一回事呢?今天的文献分享就告诉您~

发表期刊:Developmental cell
发表单位:美国杜克大学、德国柏林洪堡大学
影响因子:13.417
发表时间:2022.02
测序策略:单细胞转录组(scRNA-seq)
研究背景
多细胞生物通过控制的转录网络明确细胞特性,关联位置信息,协调器官的发育和成熟。明确这些网络如何协调器官的发育和功能,需要详细地理解该组织器官的时空基因表达模式。拟南芥的根是一个易于操作的模式器官,具有大多数细胞类型的标记,以及形态学定义的发育阶段。有利于进行器官发育过程中的基因时空表达模式的研究。尽管,此前已有一些研究单位报道拟南芥单细胞测序图谱,但是这些一代图谱中没有一个使用超过12,500个细胞。而且仅有一个研究单位推断了三种以上细胞类型的发育进程。因此,目前还没有一个全面的拟南芥根系图谱,可以捕捉到所有主要细胞类型的发育状态的细微变化。
研究材料
材料:拟南芥野生型( Col-0 ) 96,000个根细胞(13个样本),scr突变体10,000个根细胞(n=2),shr突变体10,000个根细胞(n=2)。
方法:单细胞转录组测序、碘化丙啶(PI)染色
主要结果
1. 构建拟南芥根的scRNA-seq图谱,注释细胞类型和发育阶段
为了绘制一个全面的、器官尺度的拟南芥单细胞分辨率图谱。作者选取5-7日龄的野生型幼苗主根尖0.5 cm的组织进行取样,制备原生质体,使用10X Genomics scRNA-seq平台对超过96,000个根细胞进行了分析。之后结合已发表的scRNA-seq相关数据(3个样本14,427个细胞),最终构建了单细胞水平上的十万级根细胞时空表达图谱。数据统计显示,每个细胞平均检测出2,768个基因,总计24,997个基因,占拟南芥基因组中编码基因的90%。作者推断出jing q 的细胞类型注释,将每个细胞分配到14个细胞类型和7个发育阶段。之后,作者结合novoSpaRc、SEMITONES、ICI(index of cell identity)等方法,通过每个细胞的表达谱来确定细胞类型和发育阶段之间的边界(图1、图2)。
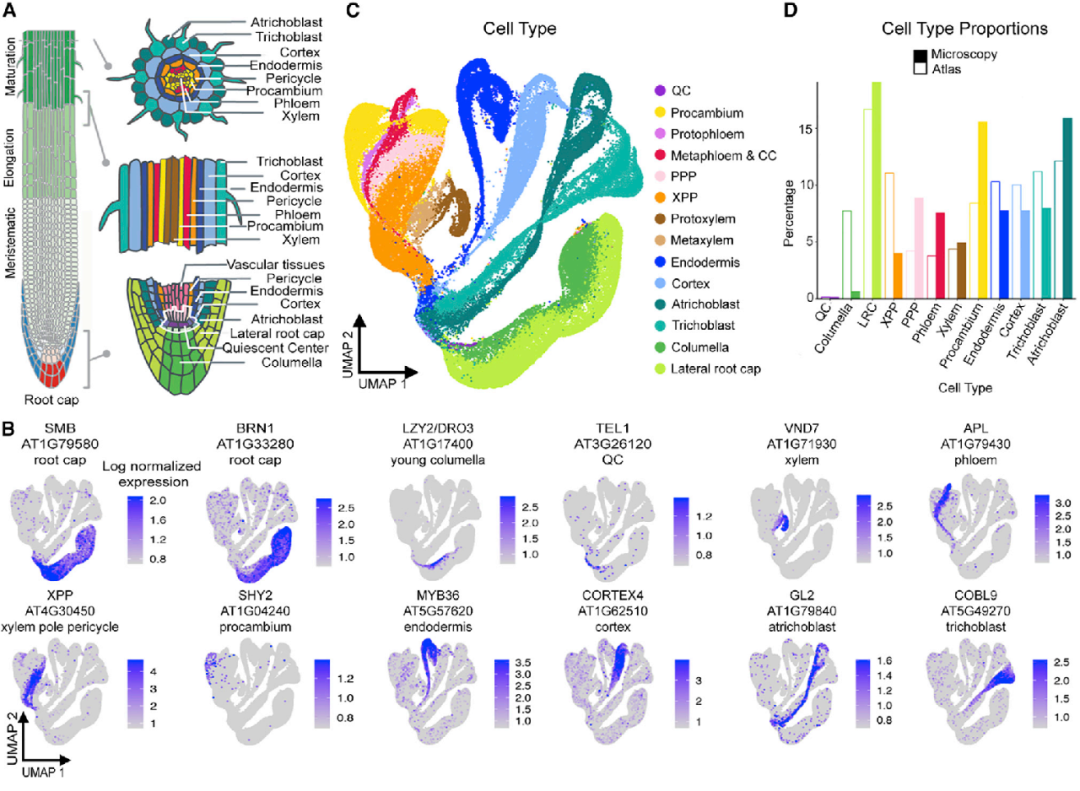
图1 代表所有主要细胞类型的110,427个细胞根部图谱
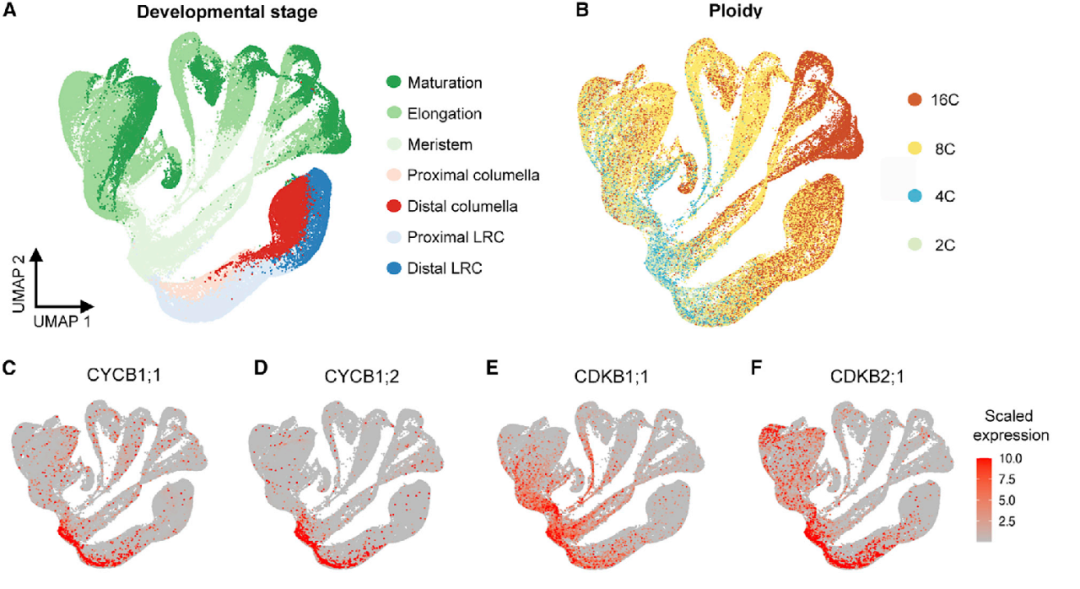
图2 通过已知基因的表达谱来辅助图谱的发育阶段注释
2. 通过各个组织类型来推断发育进程
为了更详细地分析根的各个细胞的发育进程,作者从注释的细胞系内的简单拟时序分析开始,基于干细胞的起源将图谱细分成4个组织/细胞系群。并使用两种不同方法的非图形工具CytoTRACE和scVelo来量化细胞的分化进程。通过将拟时间顺序划分为10组(T0-T9)进行差异表达分析,确定了基因的动态表达随着皮层和内皮层的分化循序渐进(图3)。同样,在中柱细胞、表皮、侧根冠细胞(LRC)和中轴细胞之间,基因也呈现出重叠渐进的表达模式。
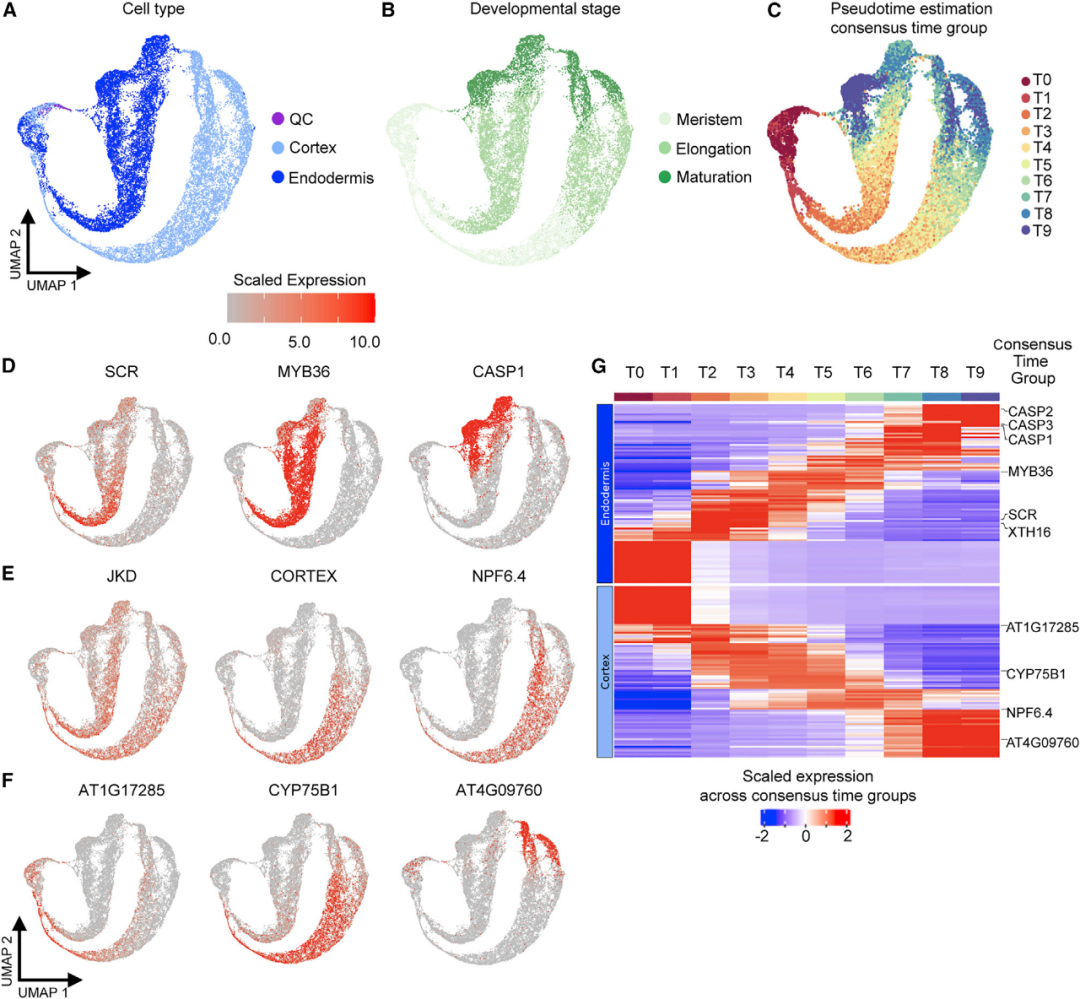
图3 拟时序估计反映了基本组织分化的动态
3.理想传输理论确定发育轨迹
由于拟时序推理表明根细胞类型以不同的速率成熟,于是作者利用基于理想传输理论OT(Optimal Transport)的StationaryOT分析方法来推断整个图谱的发育轨迹。研究人员发现它可以在很大程度上独立于图谱注释和细胞系分割,来推断细胞系的发育轨迹,反映每种细胞和组织类型分化的现有生物学知识。分化事件可以通过将多个命运概率以“三角图”的形式投影到重心坐标中来实现可视化。作者利用“三角图”分别展现了分生区、伸长区、成熟区的内皮层和皮层细胞的命运概率(图4D-F)。已知内皮层标记基因SCARECROW (SCR) 和MYB36的表达模式与伴随细胞成熟而不断增加的内皮层命运概率相一致(图4G)。之后按照发育区对生毛细胞、非生毛细胞(图4J-L),中柱鞘、原形成层(图4O-Q)也绘制“三角图”。此外,理想传输理论也有助于识别发育调节因子。
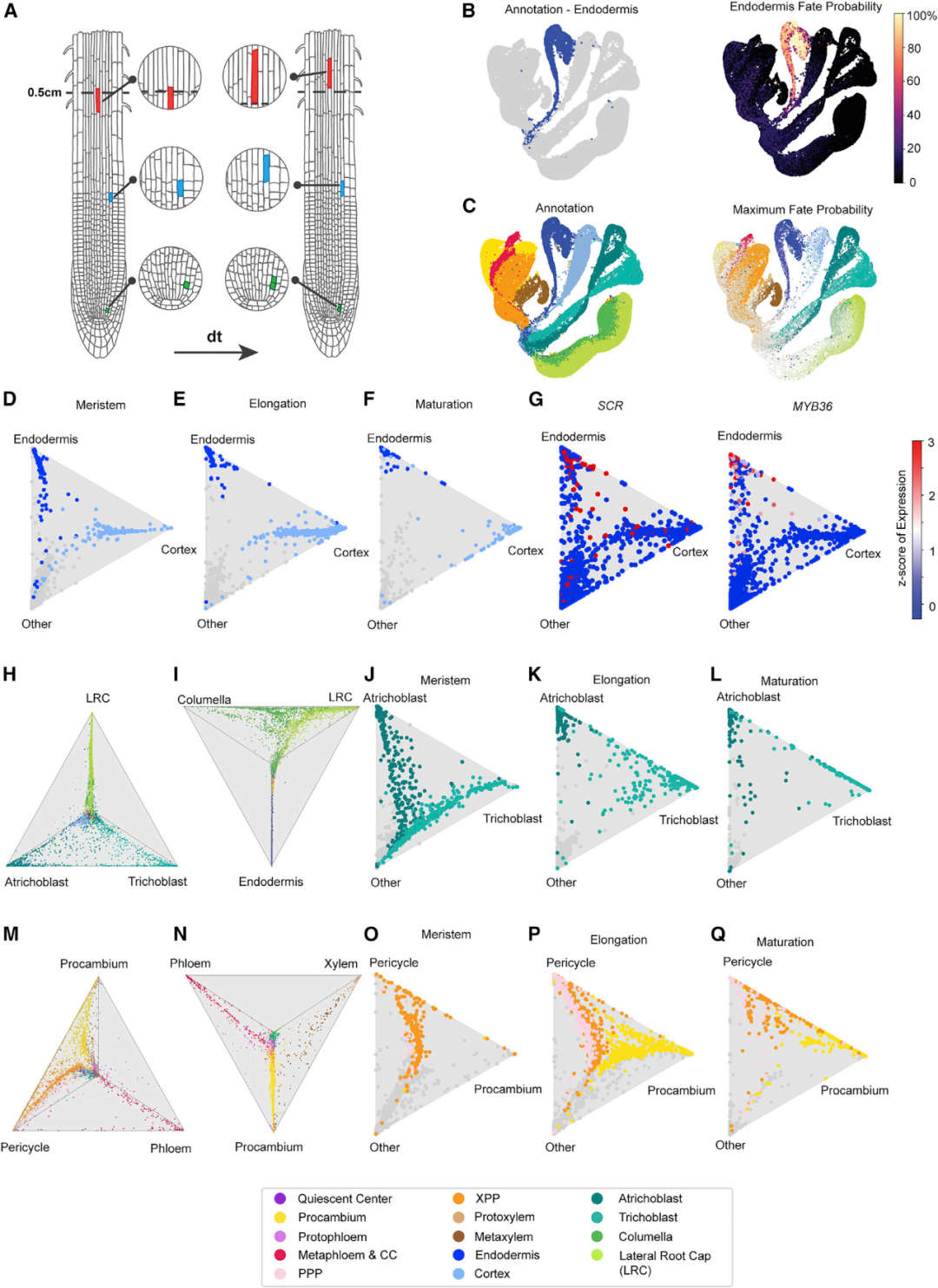
图4 理想传输法确定发育轨迹
4.scRNA-seq揭示scr突变体的分化途径
在根的发育过程中,转录因子SHORTROOT (SHR) 和SCR对干细胞龛维持和组织模式形成至关重要。那么,SHR或SCR的功能丧失会如何影响组织构成以及根细胞的特性和分化呢?由于shr和scr突变体都缺乏形成基本组织的不对称细胞分裂,导致形成了一个单一的突变体组织层,而不是皮层和内皮层细胞层。与先前组织特异性标记与形态检测的突变体表型一致,通过对shr和scr突变体细胞的scRNA-seq数据分析发现,它们中表达内皮层marker基因的细胞显著减少。两个突变体原生木质部细胞的丰度也明显下降了,并且shr中原生韧皮部和初生韧皮部细胞丰度下降,这与shr和scr中柱缺陷的表型一致。此外,该研究发现木质部和韧皮部中柱鞘細胞(pericycle cell)对应的基因表达也下调了;这一结果与shr突变体影响侧根发育一致(图5)。
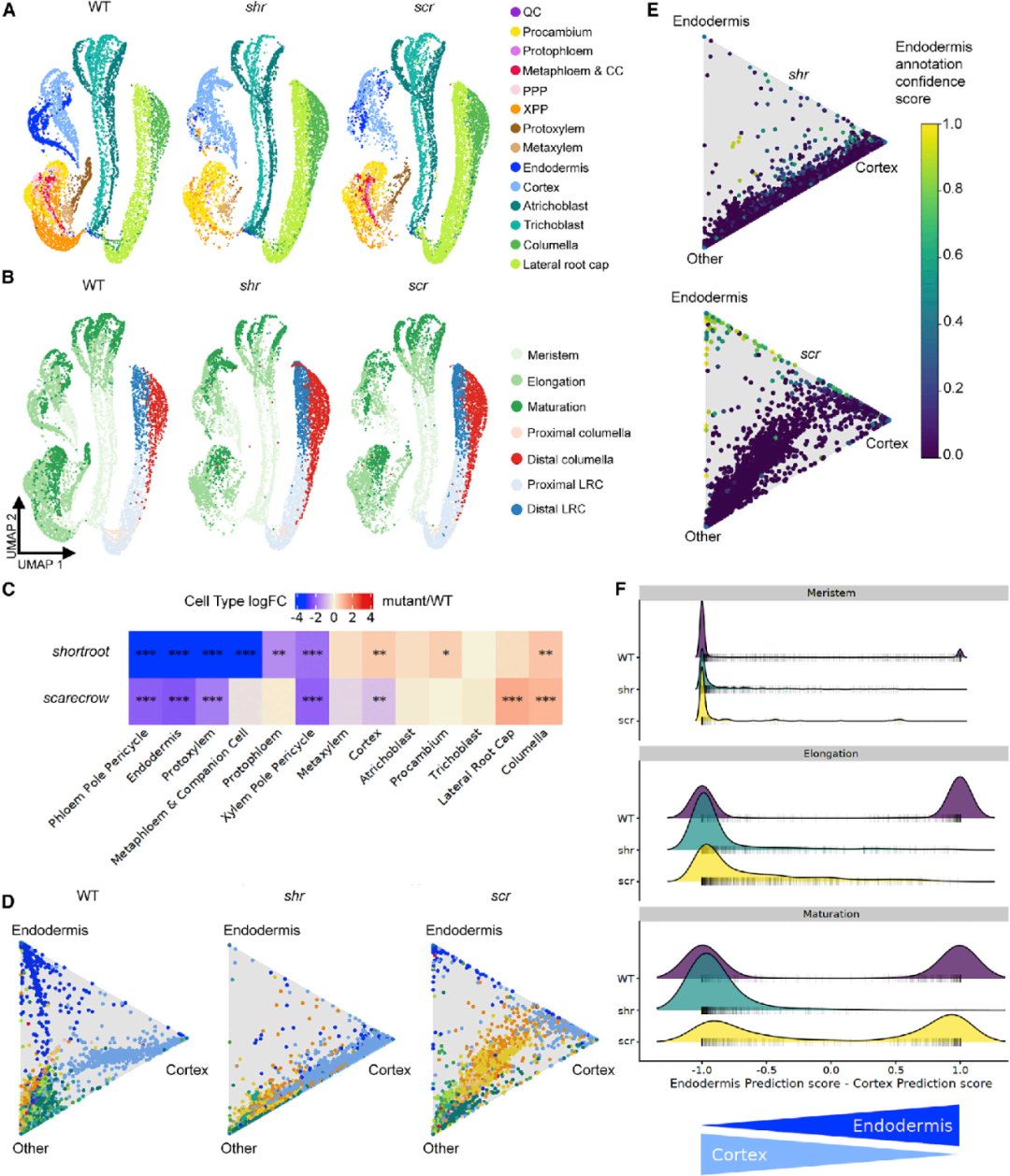
图5 图谱揭示了shr和scr突变体的细胞类型丰度和特性变化
5.scRNA-seq表明scr突变层的转分化
接下来,一个值得思考的问题是单一细胞是如何对报告的scr突变体层的混合特性做出贡献的?从单一细胞的发育轨迹来说,造成这种混合特性的可能性有多种。1)细胞在发育早期获得了内皮层和皮层的身份,而突变层是将两种细胞类型的异质性混合;2)每个细胞可能具有皮层和内皮层的混合属性;3)细胞在发育早期只获得一种特性,随后改变其命运。通过StationaryOT分析来计算scr细胞的命运概率最终发现scr突变层细胞同时具有皮层和内皮层的特性。有意思的是,scr突变层细胞在发育早期(分生区)主要是类似皮层,而成熟区的细胞呈现出内皮层的特性(图6-1)。
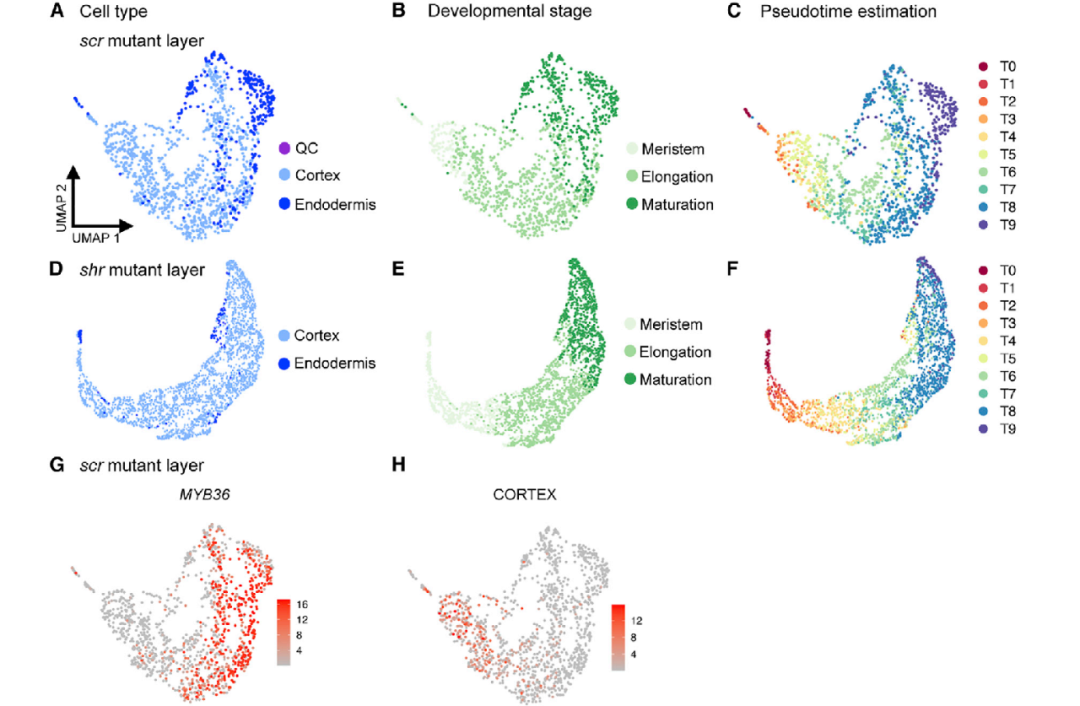
图6-1 scr突变层细胞由皮层到内皮层的特性转变
为了在体内进一步对scRNA-seq得到的结论进行验证,作者验证了皮层和内皮层标志物的空间表达模式在scr突变层是否会发生改变。在WT中,内皮层标志基因MYB36和皮层转录报告基因AT1G09750 (CORTEX)在它们各自类型的伸长区表达,MYB36在分生区的内皮层也表达。但scr突变体中,MYB36转录报告基因的表达仅在晚期延伸区和成熟区可见,皮层转录报告基因在延伸区和成熟区都有所减少(图6-2)。以上结果都与scRNA-seq分析结果一致。
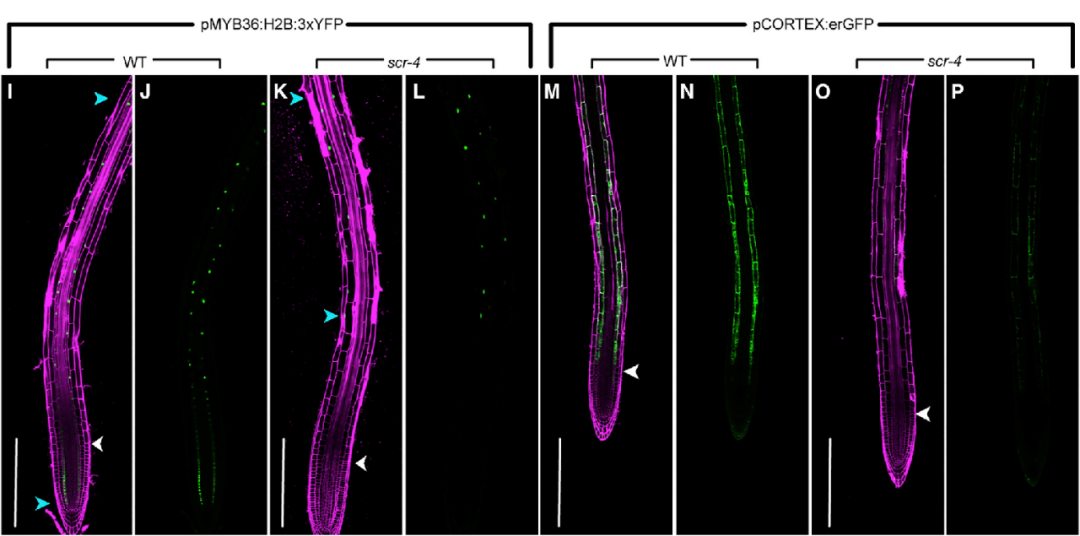
图6-2 野生型和scr突变体中MYB36和CORTEX(皮层转录报告)基因的空间表达模式
总结
这篇文章通过对不同发育阶段的拟南芥根细胞进行scRNA-seq,构建了十万级水平的根细胞基因时空表达图谱,并利用理想传输理论(OT)推断出了细胞系的发育轨迹。同时,通过比较scr和shr突变体的scRNA-seq数据,从单细胞水平阐释了scr突变层细胞同时具有皮层和内皮层混合特性,并指出了其发育轨迹上的特性转变。最后,使用内皮层和皮层转录报告基因对以上结论进行了体内验证。
参考文献:
Shahan R, Hsu CW, Nolan TM, et al. A single-cell Arabidopsis root atlas reveals developmental trajectories in wild-type and cell identity mutants. Dev Cell. 2022 Feb 28;57(4):543-560.e9.




 京公网安备 11011302003368号
京公网安备 11011302003368号