

在中心法则中,RNA处于重要的中央枢纽地位,转录组作为生信科研中的“万金油”,几乎可以与所有的组学产品联合分析。转录组代表了基因表达的中间状态,可以反映诸如转录调控、转录后调控的机理;蛋白组代表生物体直接功能执行状态,可以反应转录本真实的表达情况;而代谢组可以反映生物体表型的状态变化。转录组+蛋白组+代谢组的多组学分析可以同时实现从“因”和“果”两个层面来探究生物学问题,相互间进行验证,从海量的数据中筛选出关键基因、蛋白质及代谢通路,解释生物过程的复杂性和整体性,提高文章的水平。今天小编通过一篇案例的解析,带领大家进入多组学联合分析的新视野。
英文题目:Multiomics Analysis Reveals New Insights into the Apple Fruit Decline under High Nitrogen Conditions
中文题目:多组学联合分析揭示了高氮条件下苹果果实品质下降的新见解
发表期刊:Journal of agricultural and food chemistry
影响因子:5.279
原文链接:https://pubmed.ncbi.nlm.nih.gov/33945277/
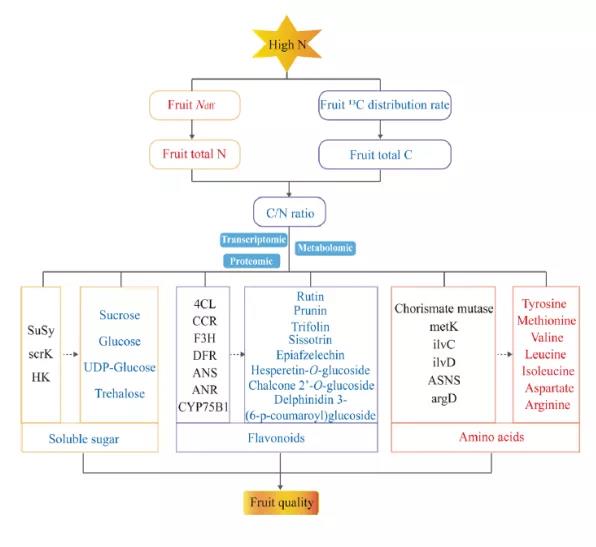
研究背景
水果质量是消费者偏好的最重要指标,包括各种质量特征,如水果颜色、甜度、风味和健康价值。水果的甜度取决于各种糖和糖醇的浓度和组成。此外,次生代谢产物,特别是黄酮类化合物,被认为是人类日常消费的重要物质,可以降低癌症和心血管疾病的风险。
在中国,过量施用氮肥已成为一种普遍现象,苹果树种植中施氮量已达600−800 kg/hm2,远远超过苹果树的需求。虽然高氮对提高产量和单果重有积极作用,但过量的氮会影响C−N平衡,对可溶性糖、类黄酮等果实品质指标产生负面影响。以往的研究主要集中在氮胁迫对苹果叶片的影响上,而高氮对苹果果实初级代谢和次级代谢影响的研究相对较少。本研究通过转录组学、蛋白质组学和广靶代谢组学的联合分析,探索过量氮营养与初级和次级代谢物积累之间的关系,特别是C-N平衡中碳水化合物、氨基酸和黄酮类化合物的变化,并为提高苹果果实品质提供新的见解。
材料方法
实验材料:红富士苹果(7龄;中国东北部山东省烟台市)。对照组CK:正常尿素处理组(300 kg N/hm2,每棵树435 g尿素);实验组MHN:中高氮处理组(600 kg N/hm2,每棵树870 g尿素);实验组HN:高氮处理组(800 kg N/hm2,每棵树1160 g尿素)。在开花后183天进行13C标记。
理化指标测定:氮含量(凯氏定氮法);15N和13C丰度(DELTAV advantage同位素比值质谱仪);Ndff和13C分布率;可溶性固形物(糖量计)和可溶性糖的含量(蒽酮比色法);总游离氨基酸(茚三酮比色法)和可溶性蛋白质含量(Bradford法)。
组学研究:RNA-seq;TMT标记定量蛋白质组;广靶代谢组
主要结果与分析
1. 生理分析
Ndff是指各器官从肥料中吸收的15N相对于各器官总N的贡献率,它反映了各器官吸收和调节15N的能力。与对照相比,MHN和HN使果实、一年生枝条和叶片的Ndff值增加,并随施氮量的增加而增加(图1A),表明了果实对15N的吸收和调节最大,MHN和HN提高了新生器官对15N的吸收和调节能力。与对照相比,MHN和HN降低了果实的13C分布率,增加了营养器官(一年生枝和叶)的13C分布率(图1B),且MHN和HN果实中15N积累增加,而13C的积累减少。因此,MHN和HN提高了果实对15N的吸收和调节能力,减少了13C从营养器官向果实的分配。另外,MHN和HN使果实可溶性糖和总黄酮含量分别降低16.05%和19.01%(图1I,J),表明高氮条件下苹果果实品质恶化。此外,与对照相比,MHN和HN增加了总游离氨基酸(图1K)和可溶性蛋白质(图1L)的含量。CK和HN果实生理指标上不存在显著差异,因此选择这两个处理的果实样品进行后续的组学研究。
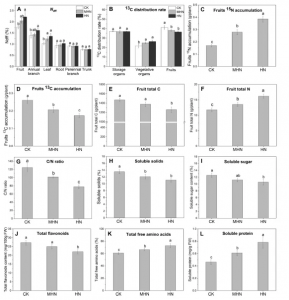
图1 苹果生理指标
2. 转录组分析
结果显示204个基因上调,545个基因下调,KEGG分析显示差异基因(DEGs)主要参与碳水化合物代谢、氨基酸代谢和其他次级代谢产物的生物合成(图2B)。此外,共鉴定出91种转录因子(TF),涵盖26个TF家族(图2C)。总的来说,大多数TF在高氮水平下下调。AP2-EREBP、MYB和bHLH是相对较大的TF家族。结果表明,这些显著变化的TFs在高氮胁迫下起着重要作用。
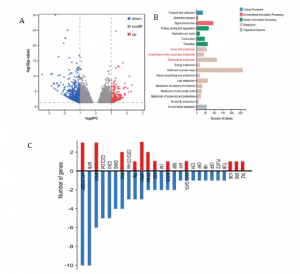
图2 转录组学分析
3. 蛋白质组分析
共获得5340个蛋白质,与对照相比,795个DEPs在HN中显示出显著变化,其中422个蛋白上调,373个蛋白下调。GO富集分析显示氧化作用−还原过程和应激反应富含更多的蛋白质,而下调的蛋白质富集程度更高(图3A)。KEGG分析发现C代谢、丙酮酸代谢、糖酵解/糖异生、三羧酸(TCA)循环、次级代谢产物生物合成和氨基酸生物合成途径显著富集(图3B)。MapMan分析结果显示许多DEP参与主要和次要CHO代谢、糖酵解、TCA循环、氨基酸代谢和次级代谢,并且大多数这些蛋白质被发现下调(图3C)。
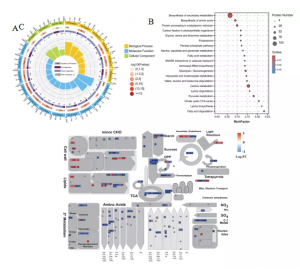
图3 蛋白组分析
4. 广靶代谢组学分析
共鉴定到36个上调代谢物和47个下调代谢产物(图4B),包括类黄酮,碳水化合物、氨基酸和衍生物等(图4C)。KEGG富集分析表明差异代谢物(DAMs)主要在氨基酸生物合成、次生代谢产物生物合成、C代谢和类黄酮生物合成富集。碳水化合物中D-果糖6-磷酸和Dglucono-1,5-内酯的下调倍数最大,氨基酸和衍生物中,有四种下调,六种上调,其中高精氨酸的上调倍数最大。此外,结果还表明果实中积累的黄酮类化合物较少,且氮含量较高,尤其是鼠李素-3-O-芦丁苷和芦丁。
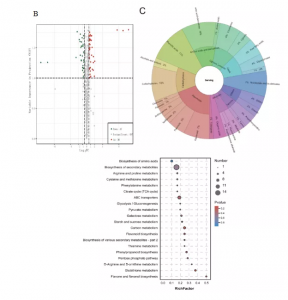
图4 HN与CK的代谢组学分析
5. 初级代谢的联合分析
C代谢包括淀粉和蔗糖代谢、糖酵解和TCA循环。高氮总体下调了C代谢中蛋白质(Susy、scrK和HK)的丰度和碳水化合物(蔗糖、葡萄糖和海藻糖)的积累。相反,糖酵解中间产物却明显积累,如甘油-P、甘油酸-3P和磷酸烯醇式丙酮酸。这表明蛋白质(ALDO、gapN和ENO)与这些中间代谢物之间存在负相关。糖酵解亚型的增加表明,高氮加速了C在呼吸途径中的流动。氨基酸与碳代谢密切相关,是氮分配的主要形式。氨基酸生物合成途径中的大多数蛋白质表达下调(脊索酸变位酶除外),而大多数氨基酸及其中间产物表达上调(图5)。以前的研究表明,随着氮供应的增加,非结构性碳水化合物中的一部分C被并入氨基酸和蛋白质中。以氨基酸形式的氮增加,使氮储存更具成本效益。因此,C向N代谢的转移是导致高氮条件下碳水化合物减少的一个重要因素,这使得水果中氨基酸和总氮含量的显著增加。甘油酸-3P和磷酸烯醇式丙酮酸的增加是氨基酸生物合成所需C骨架增加的重要指标。
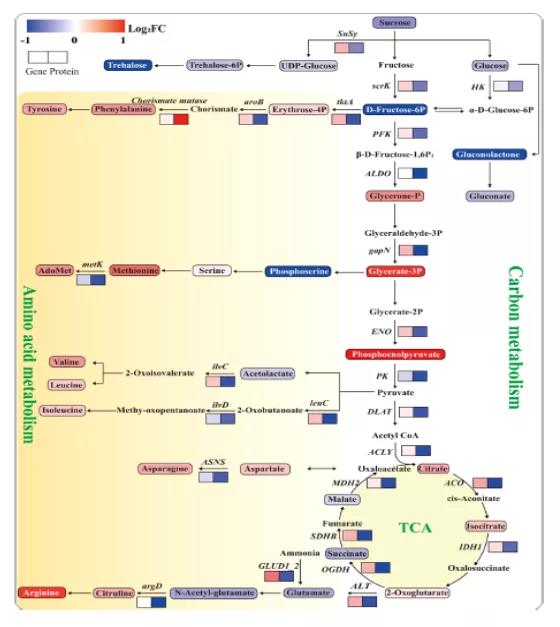
图5 C代谢和氨基酸生物合成途径
6. 次级代谢联合分析
参与苯丙烷生物合成途径的蛋白质(4CL、CCR、C3′H和UGT72E)下调,下游代谢物东莨菪碱也显示下调趋势。相反,苯丙烷代谢的另一个分支的代谢产物(松柏醛和松柏醇)显著上调,这可能是由于苯丙烷途径分支之间的代谢通量重新定向,以应对高氮胁迫并维持苯丙烷代谢的动态平衡。黄酮类化合物来源于苯丙烷代谢途径,由C代谢诱导。参与类黄酮生物合成的基因(CCR、TCEA、F3H、DFR、ANS、BZ1和CYP75B1)和蛋白质(PGT1、F3H和ANR)表达下调。类似地,参与类黄酮生物合成途径的代谢物(根皮素、剑麻素、橙皮素-7-葡萄糖苷、三叶草素、槲皮苷、芦丁等)显示出下调趋势,其中,芦丁的表达下调幅度最大(图6)。在类黄酮生物合成途径中,发现许多TF可调节结构基因的表达。此外,在高氮条件下,C在初级和次级代谢中的分布可能是类黄酮生物合成下降的原因。
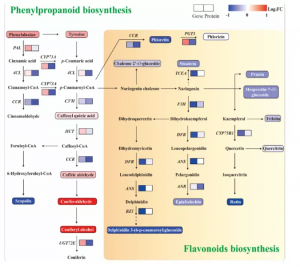 图6 苯丙素和类黄酮的生物合成途径
图6 苯丙素和类黄酮的生物合成途径
7. 共表达网络分析
基于Pearson相关分析(系数>0.9,P<0.05),筛选与初级代谢和次级代谢相关的基因、蛋白质和代谢物(图7)。获得5个hub基因和3个hub蛋白,其中MD07G1172700(编码PAL)包含在次级代谢中,MD05G1222800(编码argD)、MD16G1227200(编码leuC)、MD01G1174400(编码GLUD1_2)、MD02G1207200(编码tktA)、PFK、gapN,而HK则包含在初级代谢中。这些hub基因和蛋白质之间存在着相对较多的正相关节点。

图7 参与初级和次级代谢途径的基因、蛋白质和代谢物的共表达网络
总结
高氮增加了果实对15N的吸收和调节能力,降低了C在果实中的分配率,从而导致苹果果实的总N增加,总C和C/N比率降低。高氮破坏了苹果C−N平衡,降低了苹果果实的可溶性糖和总黄酮含量,进而使果实品质恶化。基于多组学分析,揭示了高氮对初级代谢和次级代谢互作网络。高氮调节了苹果果实中基因和蛋白质的表达,抑制了碳水化合物和黄酮类化合物的积累,更多的碳骨架被用来合成氨基酸及其衍生物并转移到氮代谢中。本研究揭示了高氮导致苹果果实糖和类黄酮含量下降的机理,并获得了关键候选基因和蛋白质。
参考文献
Wang F , Ge S , Xu X , et al. Multiomics Analysis Reveals New Insights into the Apple Fruit Quality Decline under High Nitrogen Conditions[J]. Journal of Agricultural and Food Chemistry, 2021, 69(19).




 京公网安备 11011302003368号
京公网安备 11011302003368号