

2019年03月11日,Nature Communications在线发表了西南大学农学与生物科技学院与北京百迈客生物科技有限公司共同合作文章“Whole-genome resequencing reveals Brassica napus origin and genetic loci involved in its improvement” 西南大学卢坤研究员为本文第一作者,西南大学李加纳教授、中国农业科学院蔬菜花卉研究所王晓武研究员、美国佐治亚大学Andrew H. Paterson教授为该文章共同通讯作者,其中北京百迈客公司生物信息研发总监周钢博士,王运通,李小龙为共同作者,文章详细解读如下:
英文题目:Whole-genome resequencing reveals Brassica napus origin and genetic loci involved in its improvement
中文题目:全基因组重测序揭示甘蓝型油菜起源和参与其改良的遗传基因座;
发表杂志:Nature Communications ;
影响因子:IF=12.353;
发表时间:2019.03.11;
合作单位:西南大学农学与生物科技学院;
摘 要
甘蓝型油菜(Brassica napus,2n=4x=38,AACC)是一种重要的异源多倍体作物,其来自于白菜型油菜(Brassica rapa,2n=2x=20,AA)和甘蓝(Brassica oleracea,2n=2x=18,CC)的种间杂交。然而,真正的野生B. napus群体尚不清楚,其起源和改良过程仍不清楚。在本研究中,研究人员利用重测序的方法对588份甘蓝型油菜(Brassica napus)进行测序,揭示了A亚基因组可能从欧洲芜菁的祖先进化而来,而C亚基因组可能是从大头菜,花椰菜,西兰花和中国羽衣甘蓝的共同祖先进化而来。此外,冬季油籽型可能是欧洲油菜的原始形式。防御反应相关基因在亚基因组上的特异性选择有助于物种形成后的环境适应性,而不对称的亚基因组选择导致生态型的变化。通过结合全基因组关联分析(genome-wide association studies),群体选择分析及转录组分析,研究人员鉴定了与提高的胁迫耐受性,含油量,种子质量和生态型改良相关的基因,这些基因成为甘蓝型油菜(Brassica napus)进一步功能鉴定和遗传改良的候选基因组。
研究背景
甘蓝型油菜(Brassica napus)是世界范围内重要的油籽作物,包括用于动物饲料和人类消费的结节(甘蓝或大头菜)和绿叶(饲料油菜和羽衣甘蓝)形式存在。在Brassica napus的育种期间,种子中不需要的油用成分芥酸和脂肪族硫代葡萄糖苷含量显着地减少,而含油量,种子产量和抗病性得到了显着改善。如禹氏三角(Triangle of U)中所描述,三个异源多倍体种,甘蓝型油菜Brassica napus (AACC, 2n = 38),芥菜Brassica juncea (AABB, 2n = 36)和埃塞俄比亚芥Brassica carinata (BBCC, 2n = 34) 分别来源于三个祖先二倍体种间两两杂交,三个祖先二倍体种包括:芜菁Brassica rapa (AA, 2n = 20),甘蓝Brassica oleracea (CC, 2n = 18),和黑芥Brassica nigra (BB, 2n = 16)。这三个芸薹属二倍体祖先种本身是古多倍体,经历了谱系特异的全基因组三倍化,使得异源多倍体后代甘蓝型油菜(B. napus)成为研究多倍体形态,进化和选择过程的理想模型。
作为较早的异源多倍体作物之一,B.napus通过B. rapa和B.oleracea杂交形成,估计甘蓝型油菜(B.napus)的形成时间为~6700和~7500年前及38000-51000年前。文献中记载,冬季甘蓝型油菜首次在欧洲种植,大约在1700年,春季甘蓝型油菜开始种植并在18世纪末传播到英国,半冬生态型主要在中国种植,在1930 – 1940年间从欧洲引入。然而,并没有真正已知的野生甘蓝型油菜(B.napus)群体,导致杂交形成B.napus的两个祖先种的√确身份仍然难以捉摸,因为B. rapa和B. oleracea具有形态多样的亚种,并且通常在整个欧洲种植了几百年。这些物种之间的天然杂交偶然性地在适当的条件下发生。虽然最近的一项研究表明甘蓝型油菜(B.napus)的A亚基因组可能来源于欧洲芜菁(B. rapa ssp. rapa)的祖先,但是由于仅5份B.napus和27份 B. rapa种质被纳入该项分析中,因此需要提供更多支持该结论的证据。先前基于细胞核和叶绿体标记的研究也表明,甘蓝型油菜可能是从萝卜和西兰花之间的种间杂交发育而来,或是由几个独立的杂交事件形成的。为了进一步了解甘蓝型油菜的进化,有必要揭示野生种或驯化种的供体是否是亲本祖先,其中B. rapa和B. oleracea亚种参与甘蓝型油菜的形成。
全基因组重测序这一技术手段已被用于鉴定有助于作物驯化和改良的基因,例如水稻(Oryza sativa),番茄(Solanum lycopersicum)和大豆(Glycine max)。三个芸薹属祖先种和异源多倍体种B. napus和B. juncea的基因组已经释放。199份B. rapa和119份B. oleracea种质的重测序将有助于研究B. napus中每个亚基因组中的受选择作用,同时将澄清该物质的祖先种。
在本研究中,研究人员进行了588份B. napus材料的重测序及2份具有不同种子质量的B. napus种质中11个组织的转录组测序。大量变异的鉴定不仅仅提供了对甘蓝型油菜(B. napus)起源和进化历史的深入理解,确定了其改良所涉及的遗传基因座,而且为控制重要性状的候选基因的进一步功能验证奠定了基础。此外,本研究的发现为促进B. napus和具有优良性状的相关作物的繁殖提供了基础。
材料方法
研究材料:588份甘蓝型油菜(B. napus)种质(466份来自亚洲,102份来自欧洲,13份来自北美,7份来自澳大利亚),根据生长习性记录,这些材料分为三种生态型[春季型(86份),冬季型(74份)和半冬季型(428份)];199 B. rapa和119 B. oleracea种质数据(NCBI中下载);2份具有不同含油量和种子质量的代表性种质(Zhongshuang11和Zhongyou821)的11个组织;
研究策略:Illumina HiSeq 4000,PE125,插入片段350 bp;B. rapa (version 1.5) 和B. oleracea (version 1.0)整合为甘蓝型油菜祖先的假基因组(B. napus ancestral pseudo-genome);B. napus参考基因组(version 4.1);
研究方法:
参考基因组比对:Burrows-Wheeler Aligner (version 0.7.10-r789),Picard(release 2.0.1),GATK(version 3.2-2-gec30cee);
SNP和InDel calling: SAMtools’s mpileup (version 0.1.19-44428cd)和GATK,SelectVariants in GATK(以B. napus ancestral pseudo-genome基因组为参考基因组,B. napus重测序数据进行mapping比对,calling SNPs,基于亚基因组水平,分为两组SNPs;分别以B. rapa 或B. oleracea相应的基因组做参考,在 B. rapa 或B. oleracea重测序数据中检索SNPs,与已获得的两组SNPs整合,分别代表BraA和BolC SNP集;以 B. napus基因组作为参考基因组,B. napus重测序数据进行mapping比对,获得SNPs集合Bna);
系统发育和群体结构分析:分别利用BraA, BolC和Bna SNPs数据集获得17,000, 19,377 和19,548个SNPs构建3个ML trees(IQ-TREE v1.6.6),通过Bayesian information criterion(BIC)确定*佳模型,PCA主成分分析[EIGENSOFT version 6.1.4],群体结构分析[ 50份B. napus地方种和2个祖先种,199份B. rapa和119 B. oleracea种质材料(STRUCTURE)];
B. napus的种群历史动态分析:diffusion approximations for demographic inference (∂a∂i) version 1.7.0和fastsimcoal2 v. 2.6.0.3;SMC++;
LD连锁不平衡分析: PLINK v1.07;
农艺性状的测量:本研究中所有种质材料在2013年和2014年的生长季节种植在中国重庆 (29°45′N, 106°22′E, 238.57 m) ,进一步随机选择了5株植株,进行了单株种子产量(SY),长角果长度(SL)的测量,含油量(OC),种子颜色(SC)与总硫代葡萄糖苷含量(GC)通过NIRS进行分析,脂肪酸棕榈酸,硬脂酸,油酸,亚油酸,亚麻酸,二十碳烯酸和芥酸通过气液色谱法定量,同时进行开花时间(FT)的测定;
全基因组选择清除分析:核苷酸多态性π,Fst,ROD,XP-CLR,XP-EHH;
基因型填充和z确性评估:Beagle v.4.1 ;
全基因组关联分析研究:mrMLM v1.3,GEC software;
转录组和基因本体富集分析:STAR 2.4.2a,FPKM(2个具有不同含油量和种子质量的代表性种质的11个组织,每个样品2个生物学重复);
GO富集分析:BLASTP;
研究结果
1)测序与变异分析
研究人员对来自21个国家的588份不同的甘蓝型油菜(B.napus)种质进行重测序(图1 a),共计获得4.03 Tb clean data,将测序数据过滤后,比对到甘蓝型油菜(B. napus)参考基因组上,比对效率从79.84%—99.45%不等,有效数据平均测序深度~5×(3.37×—7.71×),共计获得5,294,158个SNPs(Bna表示)和 1,307,151个indels。通过Sanger测序验证了20份种质中103个随机选择的SNPs,表明大多数已鉴定的SNPs(95.1%)是真实的。在20份种质的2个生物学重复中,对SNPs的可靠性进一步证实,发现大多数SNPs(93.5%-96.4%)是重复的。
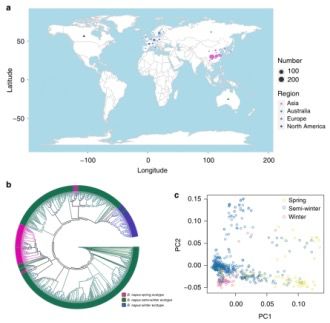
图1 甘蓝型油菜的种质的地理分布和群体结构
将B. napus测序数据比对到B. napus ancestral pseudo-genome(整合了B.rapa和B. oleracea参考基因组)上获得SNPs集合,进一步基于两个祖先种的比对信息,将SNPs数据集划分为BnaA和BnaC两个集合。将B. rapa和B. oleracea的测序数据分别比对到相应的基因组上,比对到B. rapa上的SNPs与BnaA进行整合,形成 A(B. rapa和B. napus)亚基因组SNPs集合(BraA表示,包含529,771个SNPs),类似,形成 C(B. oleracea 和B. napus) 亚基因组SNPs集合(BolC表示,包含675,457个SNPs)。
2)甘蓝型油菜(B. napus)的起源
基于*佳模型(Bna, BraA和BolC SNPs集合分别采用GTR+F+ASC+R5, GTR+F+ASC+R7 和 TVM+F+ASC+R8模型),利用IQ-TREE构建(ML) trees(图1a,2a,3a),结果显示,B. napus和B. rapa,B. oleracea各自具有一个分枝。大部分甘蓝型油菜(B.napus)依据生态型聚类到一起,而B. rapa和B. oleracea的聚类主要反映了亚种间的关系。BraA的系统发育关系显示,甘蓝型油菜(B.napus)的亲缘关系更接近于European turnip(欧洲芜菁),并与B. rapa ssp. rapa (亚洲芜菁)及其它亚种亲缘关系较远(图2a),而这正与最近的一项研究结果相一致,即甘蓝型油菜(B. napus)的A亚基因组是从欧洲芜菁的祖先进化而来的;通过动态演化情境模拟进一步分析显示,在大约~106-1,170年前,从欧洲芜菁到甘蓝型油菜A亚基因组发生了基因交流(fastsimcoal2);PCA主成分分析显示,甘蓝型油菜地方品种(B. napus landraces,n = 50,1980年前登记)更接近欧洲芜菁(n = 33)(图2b)。群体结构分析显示,当K=2时,为佳的分群数(图2c),其明确的将甘蓝型油菜的地方品种与欧洲芜菁分开,进一步支持了甘蓝型油菜A亚基因组的进化历史。
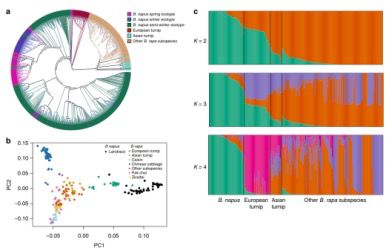
图2 588份 B. napus材料和199份B. rapa材料的群体结构分析
在BolC ML tree中,甘蓝型油菜种质(B. napus accessions)与甘蓝关系更近,主要包括4个亚种—-大头菜(B. oleracea var. gongylodes),西蓝花(B. oleracea var. italica),花椰菜(B. oleracea var. botrytis)和中国羽衣甘蓝(B. oleracea var. alboglabra),暗示了甘蓝型油菜C亚基因组可能是从这些谱系的共同祖先进化而来的(图3a)。为了验证这一结果,研究人员利用了∂a∂i和fastsimcoal2进行了动态演化情境模拟分析,∂a∂i结果支持了甘蓝型油菜C亚基因组从大头菜,西兰花和花椰菜的共同进化而来的这一模型(log-likelihood =–6806);fastsimcoal2结果以更高的支持度证明了甘蓝型油菜的C亚基因组起源于大头菜,西蓝花和花椰菜的共同祖先(Δ= 4603200; log-likelihood =–2975039),上述研究暗示甘蓝型油菜C亚基因组起源于四个甘蓝亚种的祖先分化期间;考虑到种群迁移事假可能发生在三个不同的阶段(recent, medium- and long-term),本研究中比较了五个模型(模型c-g)以推断甘蓝型油菜C亚基因组的进化历史,本研究的数据支持模型c(log-likelihood = -2914388),暗示了甘蓝型油菜的祖先从四个甘蓝亚种的共同祖先分化,甘蓝型油菜最近的基因交流发生在~108–898年前;在PCA主成分分析中,大头菜(n=19)是与甘蓝型油菜地方种亲缘关系最近的亚种,后续是中国羽衣甘蓝,西兰花和花椰菜(图3b),表明甘蓝型油菜地方品种与这四个甘蓝亚种亲缘关系更近,当K=2时,作为甘蓝型油菜地方种与甘蓝的*佳的分群数(图3c),代表不同的甘蓝型油菜和甘蓝的分类群,表明甘蓝型油菜C亚基因组具有比A亚基因组具有更复杂的起源。
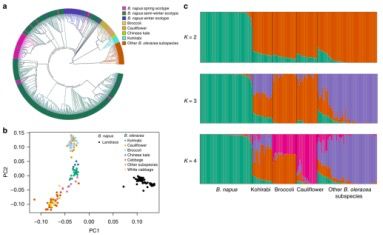
图3 588份 B. napus材料和199份B. oleracea材料的群体结构分析
Cruydt Boeck记录了冬季型甘蓝型油菜(winter B. napus)的早期形态并注意到其被用作燃料,为了进一步验证这个记录,研究人员基于A和C亚基因组的祖先种(33份欧洲芜菁和66份四种甘蓝亚种)和B. napus种质的SNP集合,比较了不同类群的LD衰减值(Linkage disequilibrium (LD) decay),在阈值r 2 = 0.3时,B.rapa(2.10 kb)和B. oleracea(27.90 kb)要强于B. napus的A(19.30 kb)和C(1365.30 kb)亚基因组,进一步支持了在甘蓝型油菜进化过程中在两个亚基因组上经历了强烈的瓶颈效应。在Bna SNP集合中,油用型甘蓝型油菜(oilseed B. napus)比菜用型甘蓝型油菜(vegetable B. napus)的LD衰减更快,冬季型比春季型和半冬季型LD衰减更快;研究人员进一步利用SMC++评估了不同生态类型的有效群体大小(Ne)和分化时间及甘蓝型油菜的用途。不同甘蓝型油菜的生态类型的Ne值显示出相似的动态,估计甘蓝型油菜的形成时间~1912-7178年前。冬季型和半冬季型甘蓝型油菜生态类型在〜60年前进行分化,而冬季型和春季型甘蓝型油菜在〜416年前进行分化,油用型和非油用型油菜在 ~277年前进行分化,而这些结果与历史记录相一致,基于上述结果,可以推测出冬季油用型油菜为原始的甘蓝型油菜。
3)改良的甘蓝型油菜受选择信号分析
首先将甘蓝型油菜起源后的改良过程分为两个阶段,第一个改良阶段(first stage of improvement,FSI)是从起源种到地方种的改良,第二个改良阶段(second stage of improvement,SSI)代表从地方种的改良到栽培种的改良。由于尚且不知道野生型甘蓝型油菜的类群,研究人员混合了两个组群的祖先种,欧洲芜菁(n = 33)和四种甘蓝的亚种(n = 66),分别以此代表了甘蓝型油菜假-野生祖先A(AA表示)亚基因组和C(CA表示)亚基因组。为了避免来自非祖先种基因渗入的影响,研究人员通过利用FST和ROD在B. napus地方种(AL表示)A亚基因组和AA之间,及在C亚基因组和B. napus地方种(CL表示)和CA之间比较基因组变异,在B. napus地方种和其祖先种中共有的变异中鉴定了FSI-受选择信号,利用甘蓝型油菜地方种作为衍生组,祖先种作为对照组。通过比较B.napus地方种和改良品种(n = 95, registration date after 2000)及不同生态类型的栽培种比较,鉴定SSI-受选择信号。
从AA到AL,核苷酸多态性(π)从7.23×10−4降到5.40×10−4,从CA到CL,核苷酸多态性(π)从7.45×10−4降到4.97×10−4,暗示了在FSI过程中,在甘蓝型油菜(B. napus)中遗传多样性丢失,且遗传多样性的丢失在C亚基因组中要强于A亚基因组,FST (AL/AA)=0.136,低于FST (CL/CA) =0.246。值得注意的是,FST (AL/AA)和FST (CL/CA) 均高于FST(improved cultivar/landrace ) (0.016),暗示了SSI过程与FSI过程相比,发生了更少的遗传分化,这种差异很可能归因于甘蓝型油菜育种史上*值得注意的事件:即广泛使用不含芥酸的品种 Liho和低硫代葡萄糖苷品种Bronowski去培育零芥酸低硫代葡萄糖苷栽培种。
通过FST和ROD的z值转换的计算检测了424和366共有FSI-选择窗口,在A亚基因组和C亚基因组上分别对应了66个和44个受选择信号,在离群区域中,研究人员分别检索了AL中的1,522个基因和CL中的811个基因。GO富集分析中揭示了FST和ROD在A亚基因上重叠的区域受选择基因主要在GO中富集在三个类群中,主要包括抗逆性[对食草动物的反应(GO:0080027),脱落酸代谢过程(GO:0009687)和免疫调控反应(GO:0050776)],生长发育[腋芽分生组织起始(GO:0090506)和分省组织起始(GO:0010014)]和代谢途径[不饱和脂肪酸生物合成过程(GO:0006636),甘油三酯生物合成过程(GO:0019432)和萜类代谢过程(GO:0006721)],而像光形态形成(GO:0009640),毛状体形态发生(GO:0010090),细胞增殖的调节(GO:0042127)和花絮的发育(GO:0010229)这些发育过程在C亚基因中过度表达。这些结果表明,A亚基因组特异性的选择可能促进了甘蓝型油菜FSI过程中耐受性和含油量的积累,而其发育特性的改善可能是由于C亚基因组中的不对称选择。关于SSI-受选择信号,研究人员通过FST , ROD, XP-CLR和XP-EHH中至少3种方法,同时鉴定了912个受选择位点(对应于跨越16.35 Mb区间内的79个受选择信号)(图4a,d)。在重叠区域鉴定的2610个基因在韧皮部硫代葡萄糖苷运输(GO:1901349)和硫代葡萄糖苷生物合成过程中(GO:0019761)过量表达——代表的相应关键基因的沉默将导致优质甘蓝型油菜种子硫代葡萄糖苷含量减少;这些基因富集到蔗糖转运(GO:0015770),RNA剪切(GO:0008380)和授粉(GO:0009856)等途径,暗示了在育种过程中除了种子质量外,还有育性,能源利用率及转录调控进行了改善。此外在双高(高芥酸和硫代葡萄糖苷)和双低(低芥酸和芥子油苷)栽培种中的84个受选择信号在1417个重叠窗口合并,并具有与SSI相似的GO富集路径。研究人员进一步确定了甘蓝型油菜(B. napus)生殖系统的的优化,受选择区的基因同时也富集到生殖过程(GO:0022414),生殖结构发育(GO:0048608)和次生枝条形成的调节(GO:2000032)这些功能。此外,参与硫代葡萄糖苷转运和脂肪酸延伸的候选基因位于A和C亚基因组的共线性区域内,这意味着平行的亚基因组选择可能有助于提高种子质量。
4)全基因组关联分析研究(Genome-wide association studies)
为了鉴定与11个农艺性状相关的候选基因,研究人员利用了多位点随机混合线性模型mrMLM v1.3进行全基因组关联分析研究(genome-wide association studies)。为了获得高质量的SNPs,研究人员对Bna SNP集合进行了估算,保留了670,028个SNP,MAF> 5%,用于后续的GWAS分析。通过19个样品多态性核苷酸和Sanger测序结果的评估比较,表明98.74%的推算基因型是正确的,20份种质的两个生物学重复推算的与真实的基因型的平均系数(r2)为0.956,最小值和最大值范围从0.928到0.967,进一步证实推算的基因型的z确性。最终共计鉴定了与10个目标性状显著相关的60个位点,其中5个与种子产量相关,3个与长角果长度相关,4个与油脂含量相关,48个与种子质量相关(图4c),此外,3个位点在阈值范围内显示与开花时间相关,与种子质量性状显著相关的SNPs解释了14.30% 到35.47%的表型变异,高于种子产量(16.81%)和含油量(20.64%),此外,本研究中还检测到与先前的GWAS研究中与种子质量相关的大部分基因座。由于与油含量和产量的相关性状是复杂的并容易受到环境变化影响,研究人员将本研究中重要的基因座位与前期研究的QTLs位点进行了整合。因此,在受选择区与QTLs定位区域内,基于功能注释和转录组分析控制不同性状的候选基因被鉴定,如下分析。
5)转录组分析(Transcriptome analysis)
为了鉴定SSI-相关基因的差异表达,研究人员收获了11个组织,从高油含量、双低栽培种Zhongshuang11和低油含量、双高栽培种Zhongyou821中选取了11个组织,过滤后,获得了848,776,815个125-bp paired-end reads,包含212.19 Gb数据量,平均82.64% 的测序reads唯一比对到甘蓝型油菜(B. napus)的参考基因组上,在开花期(Se45D)后45天,在种子中检测到了*高数量的差异表达基因(DEGs),其中包括其9,962个上调表达和12,441个下调表达基因,*低数量的差异表达基因组(DEG)发生在Se14D,仅有2,420 265个基因差异表达。
在Zhongshuang11开花后的的7天(SP7D),10天(SP7D)和14(SP7D)天,在茎(St),叶(Le)和长角果皮中下调表达的基因在芥子油苷生物合成过程中显著过量表达(GO:0019761),表明在双低品种中的低种子硫代葡萄糖苷含量可能是由硫代葡萄糖苷生物合成减少引起的。在长角果皮中,上调表达的基因在脂肪酸生物合成过程中显着过度富集,暗示着长角果皮也可能在增加种子油分含量中起重要作用。
6)甘蓝型油菜FSI期间环境适应性主要基因
在FSI时期,防御反应基因在甘蓝型油菜环境适应过程中起到了关键作用,在A亚基因组上,研究人员分别鉴定了6个,5个,6个和14个与耐旱性,食草动物抗性,对机械刺激的反应和免疫反应相关基因。在这些与耐旱相关基因中,NCED3 (Bra001552)基因在调节ABA(脱落酸)生物合成中起主要作用,以进一步响应水分缺失,除此之外,NCED3一起参与了ABA的产生,从而影响植物生长和水分胁迫耐受性;在干旱期间,内源性ABA快速生产,诱导气孔关闭,从而提高耐旱性,在A亚基因组ABA生物合成途径相关基因可能在甘蓝型油菜FSI过程中提高其干旱耐受性发挥一定的作用,为其在多样化的环境中的培育奠定了坚实的基础。其余参与ABA代谢调控的候选基因也同样值得关注,例如CYP707A3(Bra021965和Bra025083),XERICO (Bra013211)和 PHYB(Bra001650),所有这些基因都通过ABA的调节积累,与甘蓝型油菜相应耐旱机制相关。
此外,在甘蓝型油菜FSI过程中,A亚基因组的特异性选择有助于提高其抗病性,在受选择的区域,NPR3 (Bra025093)是一个值得关注的基因,作为植物免疫信号水杨酸的受体,NPR3控制蛋白酶体介导的NPR1降解,其涉及对细菌和卵菌病原体的防御反应的负调节;类似的防御反应基因,BAH1(Bra032581),NHL25 (Bra028103)和NDR1 (Bra035766)也同样参与了植物对微生物的先天免疫,进一步对甘蓝型油菜FSI过程中响应生物胁迫具有重要作用。
7)提高种子质量的主要基因
在甘蓝型油菜中,种子油分包含50%的芥酸和高水平的硫代葡萄糖苷,显着降低了其营养价值,GWAS相关结果(图4c,f)获得了与种子芥酸和硫代葡萄糖苷含量相关的几个候选基因。芥子油苷在莲座叶和长角果壁中合成,随后在转运蛋白的作用下,通过韧皮部重新定位到胚胎,研究人员鉴定了硫代葡萄糖苷特异性转运蛋白GTR2,其可能负责将硫代葡萄糖苷从源组织递送到甘蓝型油菜中的种子中,在8个Bna.GTR2基因中,3个基因(BnaC02g42280D,BnaA09g06180D和BnaA09g06190D)在双低栽培种的种子中沉默(图c,f);其中又有3个基因(BnaA02g33530D,BnaC02g42260D和BnaC09g05810D)定位到主效QTL区域中,通过沉默这5种GTR2基因,尤其是位于主效QTL区域内的基因,可以进一步减少甘蓝型油菜种子中的抗营养芥子油苷含量。在SSI期间,鉴定了4个GWAS信号与芥酸相关,包括在活跃的FATTY ACID ELONGASE 1 (FAE1) 基因[其产品控制芥酸油酰辅酶A(C18:1-CoA)合成]附近,坐落在A08 (BnaA08g11130D)和C03(BnaC03g65980D)两条染色体上的2个位点(图4)。这两个基因在芥酸品种中高量表达,表明FAE1沉默是减少种子芥酸含量的重要因素;与芥酸相关的额外的GWAS信号坐落在C02和C07染色体上(图4),在AMP-DS (BnaC02g42510D)和ATGLL (BnaC07g30920D)附近,其可能代表SSI受选择基因,其产物减少了种子中芥酸含量。
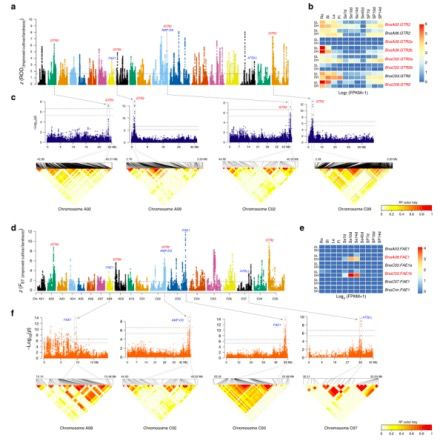
图4 甘蓝型油菜SSI期全基因组关联分析与受选择区域的注释
8)甘蓝型油菜与生态型相关基因的改良
根据春化需求,甘蓝型油菜分为冬季(需要长时间冷期),半冬季(需要短期冷期)和春季(无需冷期)共计3种生态型,*原始的甘蓝型油菜是冬季油籽型,春季型和半冬季型以适应不同的环境而发展起来。为了鉴定不同生态型下的候选基因,研究人员利用冬季型油菜作为对照,春季型和半冬季生态型作为衍生群体(图5a),在冬季型和半冬季型及冬季型和春季型油菜受选择区域,分别检测到1,996和1,117个重叠窗口,包括156个选择区域中的4,548个基因和107个选择区域中的2,729个基因,其*主要的受选择区域坐落在C亚基因组上,只有32和21个受选择区域分布在A亚基因组上,表明生态型改善主要是由不对称的亚基因组选择引起的。此外,对应于72个受选择区域的844个离群窗口在两种生态型改良分析中重叠,这些平行选择信号可能有助于甘蓝型油菜的局部适应。生态型改良选择区域的相关基因在维持花器官特征(GO:0048497),花器官脱落(GO:0010227)和花分生组织生长的调节(GO:0010080)等相关途径中具有高度代表性,暗示SSI选择信号可能对甘蓝型油菜地方环境适应性至关重要。
研究人员在FLOR-ID(FLOweRing Interactive Database)中检索到了306个基因与拟南芥开花时间相关,在甘蓝型油菜中鉴定了1,225个直系同源基因。在两组比对中,24 个开花时间相关基因同时坐落在受选择区域中(图5a,c),其中22个分布在C亚基因组上,主要通过春化和光周期途径调节开花时间。特别地,FLC基因座及其染色质介导的调节原件将是调控甘蓝型油菜开花时间的*有希望的靶标。
在春化途径中,FLC (BnaC02g00490D) 编码甘蓝型油菜生态型改良的核心调控因子(图5c),由于同源拟南芥FLOWERING LOCUS C(FLC)的产物是开花过渡期的重要阻遏物。最近的一项研究报道指出,不同的FLC旁系同源物对甘蓝型油菜开花时间的自然变异有不同的贡献,并且BnFLC.A2中的2.833-kb插入及其与BnFLC.C2的同源交换(HE)产生了早期开花的甘蓝型油菜生态型。通过GWAS研究结果显示,检测到了3个基因座位与开花时间相关(图5b),有意思的是,在染色体A02和C02上的两个FLC基因,位于2个LD blocks内(hFT-1和hFT-2),控制了开花时间的变异,解释了总计表型变异的~8%,这些结果进一步证实了生态型改良过程中FLC的重要性。通过调节FLC的表达,参与染色质修饰的基因也经历了生态型改良的广泛选择(图5a,c),例如FLC激活基因SSRP1(BnaC02g37430D),编码促进染色质转录(FACT)复合物的组分,和FLC阻遏基因VRN2(BnaC01g21540D),编码多梳抑制复合物2(PRC2)的组分。
在拟南芥中,FACT复合物中的组蛋白伴侣SSRP1有助于RNA聚合酶II的合成并促进FLC表达。相比之下,VRN2通过调节FLC基因座上H3K27me3和H3K9me3抑制标记的沉积,在春化后维持FLC的转录抑制中起关键作用。这些结果表明表观遗传变异,特别是涉及调控FLC表达的组蛋白修饰,与甘蓝型油菜的开花时间密切相关。因此,这些基因代表了未来甘蓝型油菜育种计划中有潜力的调控目标。
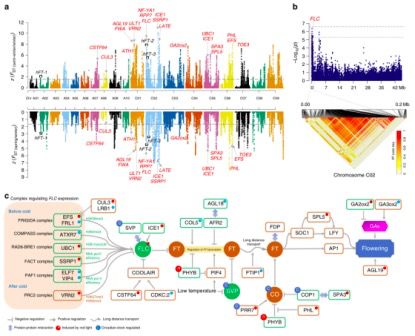
图5 受选择生态型改良甘蓝型油菜开花期调控概述
9)油籽油份含量与油籽产量相关的主要基因
种子的油份含量在甘蓝型油菜FSI期间就可能得到了显著的改善。针对油份含量,研究人员检测到了9个与QTLs区域重叠的FSI受选择区域,在这些重叠的区域中,2个基因参与脂肪酸合成,1个基因参与三酰基甘油生物合成,7个基因参与脂肪酸延伸。例如,染色体A02上的(KASI)编码可能对脂肪酸合成至关重要的蛋白质,并且还在叶绿体裂解和胚胎发育中起作用。拟南芥的KASI突变体破坏胚胎发育并显着降低种子中的脂肪酸水平,Bna.FATA1(BnaA03g37700D)是拟南芥FATA ACYL-ACP THIOESTERASE 1的同源基因,其产物通过水解酰基-ACP复合物控制末端脂肪酸合成的终止,两个拟南芥FATA基因的突变降低了FATA活性,影响油含量和种子脂肪酸组成。
甘蓝型油菜种子产量受长角果长度和枝条分枝的影响,研究人员检测到三个与长角果长度显着相关的基因座,显著的SNP-性状关联的峰值位于染色体A09上27.99 Mb,在ARF18附近的165 bp缺失,与种子重量和长角果长度增加相关;研究人员在双低甘蓝型油菜栽培种中鉴定了两对平行的SSI选择信号,拟南芥BRC1在染色体A08和C01上的直系同源基因以及DOF4.4在染色体A08和C03上的直系同源基因可能是编码甘蓝型油菜芽分枝和长角果发育的关键调控因子。因此,SSI选择信号中与种子油含量和种子产量相关的基因以及GWAS鉴定的候选基因在甘蓝型油菜育种中具有实际应用,以获得高产量和含油量的品种。
总结
本研究为油菜的起源和改良历史提供了非常重要的资源,同时有助于挖掘重要农艺复杂性状的遗传基础,其中和有利突变,选择信号和候选基因相关的显著SNP在异源多倍体油菜及其亲属的产量、种子质量、含油量和适应性上提供了丰富的宝贵资源。
小编寄语
如果您的项目有任何问题,欢迎点击下方按钮联系我们,我们将免费为您设计文章方案。





 京公网安备 11011302003368号
京公网安备 11011302003368号