

近日,河南农业大学范国强教授团队在国际期刊《Molecular Plant》(IF:13.162)上发表了题为 “Genomic insights into the fast growth of paulownias and the formation of Paulownia witches’ broom”的研究论文。百迈客生物科技作为共同一作共同参与研究。本研究通过三代PacBio等技术绘制白花泡桐基因组精细图及泡桐丛枝植原体基因组完成图,并揭示了白花泡桐速生的关键因素,以及泡桐丛枝病发生的分子机制。
研究背景
泡桐(2n=2x=40)是我国重要的速生用材树种之一,在保障我国木材安全、改善生态环境和提高人民生活水平等方面具有重要作用。然而,泡桐的利用和研究还很薄弱。泡桐丛枝(Paulownia witches’ broom;PaWB)植原体是一种专性寄生病原菌,是泡桐属植物的一种常见病害。植原体感染的泡桐,通常表现出叶片泛黄、植株生长迟缓以及丛枝等典型症状。泡桐及其植原体基因组信息的缺乏严重阻碍了PaWB病理生物学的阐明。因此,解析其对应参考基因组信息将有助于阐明丛枝病的发病机制。
材料与方法
Denovo:
1、白花泡桐(Paulownia fortunei)叶片;89X Illumina+ 87.3X PacBio+ 90X Hi-C
2、PaWB侵染的百花泡桐幼苗(PFI)的茎进行DNA分离和PacBio测序,选取无法比对到宿主基因组的Reads用于组装(泡桐丛枝植原体基因组)
RNA-seq:
1、4年生白花泡桐和毛泡桐树的叶片、树干形成层和树干韧皮部组织,每个三生物学重复。
2、健康幼苗、患病幼苗、MMS或RIF处理5天、20天等天数的患病幼苗顶芽,每个三生物学重复。
主要研究内容
1、白花泡桐基因组组装注释
2、白花泡桐系统进化与全基因组复制
3、白花泡桐形态发生相关基因家族
4、C3光合作用与景天酸代谢(CAM)相结合提高白花泡桐的光合效率
5、与泡桐植原体侵染相关的泡桐基因网络模块
6、PaWB植原体基因组特征及比较基因组分析
7、PaWB-SAP54是PaWB植原体的功能效应子
8、PaWB-SAP54通过介导PfSPLa泛素化诱导PaWB的形成
主要研究结果
- 白花泡桐基因组组装注释
本次研究的白花泡桐拥有2.03%的高度杂合,以及50%的重复序列;作者通过三代长读长等测序技术,最终组装完成511.6 Mb泡桐基因组(流式预估528.24 Mb,survey 552 Mb),contigN50=852.4 Kb;并通过Hi-C技术将93.2%序列锚定至20条假染色体上。通过BUSCO评估(97.45%)、转录组回比评估(96.02%)、Hi-C热图评估等结果表明本次构建了高质量染色体水平的白花泡桐基因组。
共预测31,985个蛋白编码基因,重复元件(257.65 Mb)占基因组的50.34%,以Ty3/gypsy和Ty1/copia的长末端重复反转录转座子(LTR-RTs)为主,分别占基因组的15.96%和13.54%。重复序列动态分析表明,在过去的800万年中,泡桐没有发生LTR-RT的爆发事件。转座子(Transposable elements,TEs)在基因组中分布不均,通常倾向于在着丝粒区积累。对于17、18号染色体,TEs在一端聚集,形成端着丝粒染色体(图1A),这与先前基于核型分析的研究结果一致。
- 白花泡桐系统进化与全基因组复制
为了进一步明确泡桐的系统进化关系,通过与透骨草科、列当科、茄科、茜草科以及葡萄科中9个近缘材料进行比较基因组分析。结果表明,泡桐科在40.92百万年时从透骨草科和列当科最近的共同祖先分化而来。在泡桐的多拷贝基因家族中,有3022个家族发生了扩张或收缩,而扩张的基因家族涉及与木质素和纤维素合成(UDP- forming)相关的基因家族。
Ks及共线性等分析表明,白花泡桐与沟酸浆(Mimulus guttatus)及芝麻(Sesamum indicum)一致,都发生过两轮全基因组复制事件。第一轮为共有的γ三倍化事件(约122~164mya的WGT-γ),第二轮为泡桐与芝麻共有的近期二倍化事件。
染色体重建表明,Chr7和Chr11是在最近的WGD之后从染色体融合中衍生出来的,因为Chr17、Chr15和Chr19分别只与葡萄Chr1、Chr18和Chr5保持了祖先的共线性(图1F)。多对泡桐染色体表现出相同的融合模式,表明染色体融合发生在最近的WGD之前:Chr8和Chr14均起源于祖先葡萄Chr9和Chr17的融合,而Chr9和Chr16均起源于祖先葡萄Chr10、Chr12和Chr19的融合。功能富集分析表明,新WGD的基因富集于“镉离子反应”和“细菌防御”,串联重复的基因富集于“生物刺激反应”和“防御反应”。这些观察结果暗示了基因组/基因复制在泡桐适应环境变化中的重要作用。

图1 白花泡桐基因组进化分析
- 白花泡桐形态发生相关基因家族
植物的形态发生和生长速率可能归因于细胞增殖和分裂、细胞壁形成以及光合效率或其他生物过程的增强。细胞分裂素生物合成中的四个基因家族和细胞周期调控中的十个基因家族,通过串联基因复制或最近的WGD发生显著扩增(图2A)。在泡桐属植物中,白花泡桐(PF)的生长速度远远快于毛泡桐(P. tomentosa, PT)。约85%的毛泡桐基因cDNA可定位于白花泡桐基因组,因此对白花泡桐和毛泡桐的叶片、树干韧皮部和树干形成层进行了比较转录组学分析。WGCNA分析得到12个潜在快速生长相关ME(FME1~12),FME3与毛泡桐和白花泡桐形成层的生长发育相关(图2B)。FME3模块包括42个参与植物激素途径或参与细胞周期调控的基因。FME4仅在白花泡桐中与形成层生长和发育高度相关(图2B),这个模块中有五个基因与激素信号有关,其中包括参与细胞分裂素代谢的腺苷激酶1,且这些基因在白花泡桐中显著扩增。通过基因的表达及拷贝数分析表明,CESAs、Csls和木质素生物合成基因可能在白花泡桐的发生中起作用(图2D)。
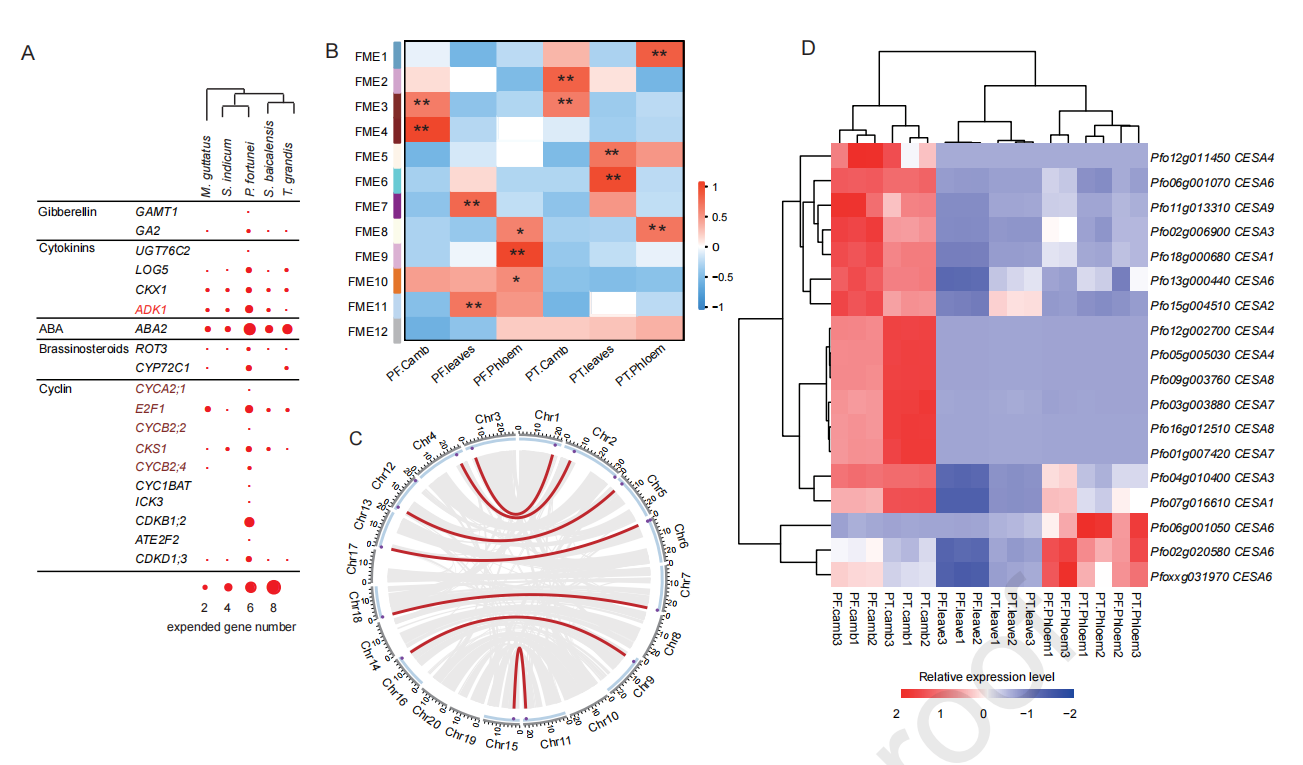
图2 参与泡桐形态发生的基因
- C3光合作用与景天酸代谢(CAM)相结合提高白花泡桐的光合效率
较高的光合效率有助于白花泡桐的快速生长。在随机选择的C3植物中,白花泡桐的净光合速率最高且与C4植物非常接近。通过Drop-seq分析白花泡桐叶肉细胞的转录组数据,发现参与卡尔文循环的基因表达水平较高,表明卡尔文循环非常活跃。此外,白花泡桐叶片的碳同位素(δ13C)变化率昼夜差异不显著。这些观察结果为白花泡桐中典型的C3碳同化提供了有力的证据。
作者测试了白花泡桐是否通过CAM来固定CO2(CAM可以通过补充C3光合作用来提高光合速率)。发现白花泡桐叶片气孔在白天和晚上均开放(图3A),夜间气孔开放表明CAM通路可能参与了泡桐的二氧化碳固定。较高的CAM途径关键基因拷贝数为泡桐中更复杂的光合类型奠定了基础。
为了确定CAM通路关键基因的表达模式,作者使用上午9:00和晚上9:00采集的白花泡桐叶片进行转录组学分析。结果表明白花泡桐通过在夜间补充CAM途径获得更高的光合能力,从而有助于泡桐的快速生长(图3D、3E)。

图3 白花泡桐CAM光合作用途径
- 与泡桐植原体侵染相关的泡桐基因网络模块
PaWB植原体感染是泡桐死亡的主要原因之一。作者采用不同浓度的甲磺酸甲酯(MMS)和利福平(RIF)处理PaWB植原体感染的白花泡桐苗(PFI)。对MMS和RIF处理不同时间的患病泡桐幼苗进行RNA-seq分析,研究泡桐对PaWB植原体侵染的响应。WGCNA分析鉴定出27个不同的PaWB相关模块特征基因(PME1~27)(图4A)。这些模块包含参与光合作用、细胞运输和离子稳态、植物生长发育、防御、以及转录有关的hub基因(图4B ,4C)。

图4 PaWB病相关的基因共表达网络分析
- PaWB植原体基因组特征及比较基因组分析
为了进一步了解植原体与泡桐相互作用的分子机制,作者对PaWB植原体基因组进行了测序和组装。组装的丛枝植原体核基因组大小为891,641 bp,有1,147个开放阅读框、32个tRNAs和4个可移动元件(PMUs),PMUs在植原体基因组的重组和植原体在宿主体内的适应性中起着重要的作用。
与另外七个植原体基因组进行比较(来自基于16srRNA的系统发育树中的I、V、X和XII分支)。通过单拷贝核心基因构建进化树,发现进化关系与16srRNA的结果基本一致, PaWB植原体与洋葱黄化植原体(OY-M)的关系更为密切(图5C)。随着系统发育距离的增加,PaWB植原体与其他植原体之间的基因组同源性大大降低,表明基因组重排的频率很高。
为了深入了解对植原体感染非常重要的代谢途径的差异,作者分析了甘油磷脂、糖酵解、嘧啶和叶酸代谢中基因的存在和缺失。只有PaWB植原体和OY-M具有完整的叶酸生物合成途径,而其他植原体缺乏这四个基因的部分或全部。

图5 PaWB植原体基因组特征
- PaWB-SAP54是PaWB植原体的功能效应子
泡桐丛枝植原体含有73个效应因子,其中11个效应因子可能与丛枝病的发生密切相关。为了研究PaWB-SAP54在PaWB植原体侵染过程中的作用,作者利用根癌农杆菌将PaWB-SAP54基因导入毛果杨。与野生型相比,转基因毛果杨表现出分枝增多,且PaWB-SAP54的转录水平增加了4.7-5.3倍(图6A)。转基因毛果杨在较细的茎上有二级分枝,而在野生型中没有观察到分枝(图6B)。表明PaWB-SAP54介导了丛枝症状的形成。
- PaWB-SAP54通过介导PfSPLa泛素化诱导PaWB的形成
为了探讨PaWB-SAP54在PaWB形成过程中的分子机制,作者采用酵母双杂交(Y2H)技术筛选了PaWB-SAP54蛋白的潜在结合蛋白。共发现112个泡桐蛋白与PaWB-SAP54相互作用,其中鉴定出的鳞状启动子结合蛋白a(PfSPLa)(图6C)是AtSPL9和AtSPL15的同源物,这两种蛋白都对腋芽的形成和分枝发育有负调控作用。在PaWB植原体感染的幼苗中,PfSPLa的蛋白质丰度显著降低(图6D)。由于泛素化参与了丛枝病的形成,因此作者研究了PfSPLa是否直接受泛素降解途径的调控。用26S蛋白酶体抑制剂环氧霉素处理感染的幼苗后,PfSPLa的表达增加(图6D),并通过westernblot分析直接证实PfSPLa的泛素化(图6E)。以PfSPLa蛋白为诱饵的Y2H筛选表明,PfSPLa可直接与多聚泛素和26S蛋白酶体非ATP酶调节亚基3(PfRPN3)相互作用(图6F)。双分子荧光互补和免疫共沉淀结果,进一步证实这种相互作用。用PfSPLa-RNAi构建转基因毛果杨,发现3个与PfSPLa同源的基因表达下调,也引起毛果杨分枝表型的发生(图6G-6I)。并且RNAi植物表现出矮化表型(图6H)。这些结果表明,PaWB-SAP54可能与PfSPLa相互作用,进而以泛素/26S蛋白酶体依赖的方式促进PfSPLa的降解,最终诱发泡桐丛枝病。
植物激素的定量显示,转基因植株中赤霉素、生长素和细胞分裂素含量有显著变化(图6J)。结果表明,PfSPLa通过控制激素水平来调节分枝的形成。

图6 泡桐丛枝植原体效应因子PaWB-SAP54与PfSPLa互作诱导白花泡桐丛枝症状形成
小结
本研究通过白花泡桐及其丛枝植原体基因组测序和进化分析,揭示了C3光合作用与景天酸代谢相结合提高光合效率的方式,是白花泡桐快速生长的关键因素。并通过WGCNA、转基因、酵母双杂交、双分子荧光互补和免疫共沉淀等方式,阐明了泡桐丛枝植原体效应因子PaWB-SAP54介导的PfSPLa泛素化引起了泡桐丛枝病的形成。该研究为加速泡桐的分子育种、品种改良及进一步阐明泡桐丛枝病发生分子机制奠定了坚实基础。





 京公网安备 11011302003368号
京公网安备 11011302003368号